Guilherme Hermanson
Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, Brazil.
Email: guilhermehermanson (at) gmail (dot) com
https://doi.org/10.5281/zenodo.10644545
Despite their secrecy, fantastic beasts are oftentimes noticed by muggles. Their diversity, however, was never subject of any study in order to understand what could have driven it. At least two groups of creatures show that both historical and environmental aspects played role on these organisms’ lineages’ splitting events, leading to their current distribution on the globe. Additionally, nonspecialist readers that enjoy Harry Potter culture might become interested in the topic and, as such, fictional content can represent an innovative tool of science outreach to introduce evolutionary biology and biogeography concepts to the general public.
WHERE AND WHO ARE THEY?
Not even special clauses (Scamander, 2001) prevented muggles of noticing fantastic beasts among them. They are part of our days probably since before we started creating tales about them (d’Huy, 2013). Present in all continents, except Antarctica, magical creatures occupy an unequal variety of niches, from herbivorous forms to fire-eating beasts (Scamander, 2001). All the main differences described for such creatures may reflect not just local traditions or modifications from oral stories, but actual lineage branching events (e.g., Hamilton et al., 2015).
Among such beasts, there are some groups in which well-known diversification processes can be exemplified, namely a clade of hominoid-related beasts, and a clade of insect-related creatures, both currently spread in the European and North American continents (Fig. 1; topology follows Gerelle et al., 2016). Basically, the appearance of natural barriers, as well as opportunistic exploitation of diverse ecological niches, could be the main causes explaining where such fantastic creatures currently inhabit (i.e., their geographic distribution); this would be in spite of the common explanation of climate change driving biodiversity dynamics (e.g., Janis, 1993; Alroy et al., 2000).
INSECT-RELATED HUMANOID BEASTS
According to Gerelle et al. (2016), Fairies, Imps, Pixies, Grindylows, and Doxies form the sister clade to butterflies (crown-Lepidoptera), making them a sort of ‘lepidopteran-like’ beasts. Despite being phylogenetically related to insects, all creatures in this clade possess humanoid traits, consisting of a remarkable case of evolutionary convergence. In addition, the absence of wings in Grindylows and Imps is probably a case of reversion to the apterous plesiomorphic condition of insects (i.e., the insect lineage was originally wingless; Kukalová-Peck, 1991).
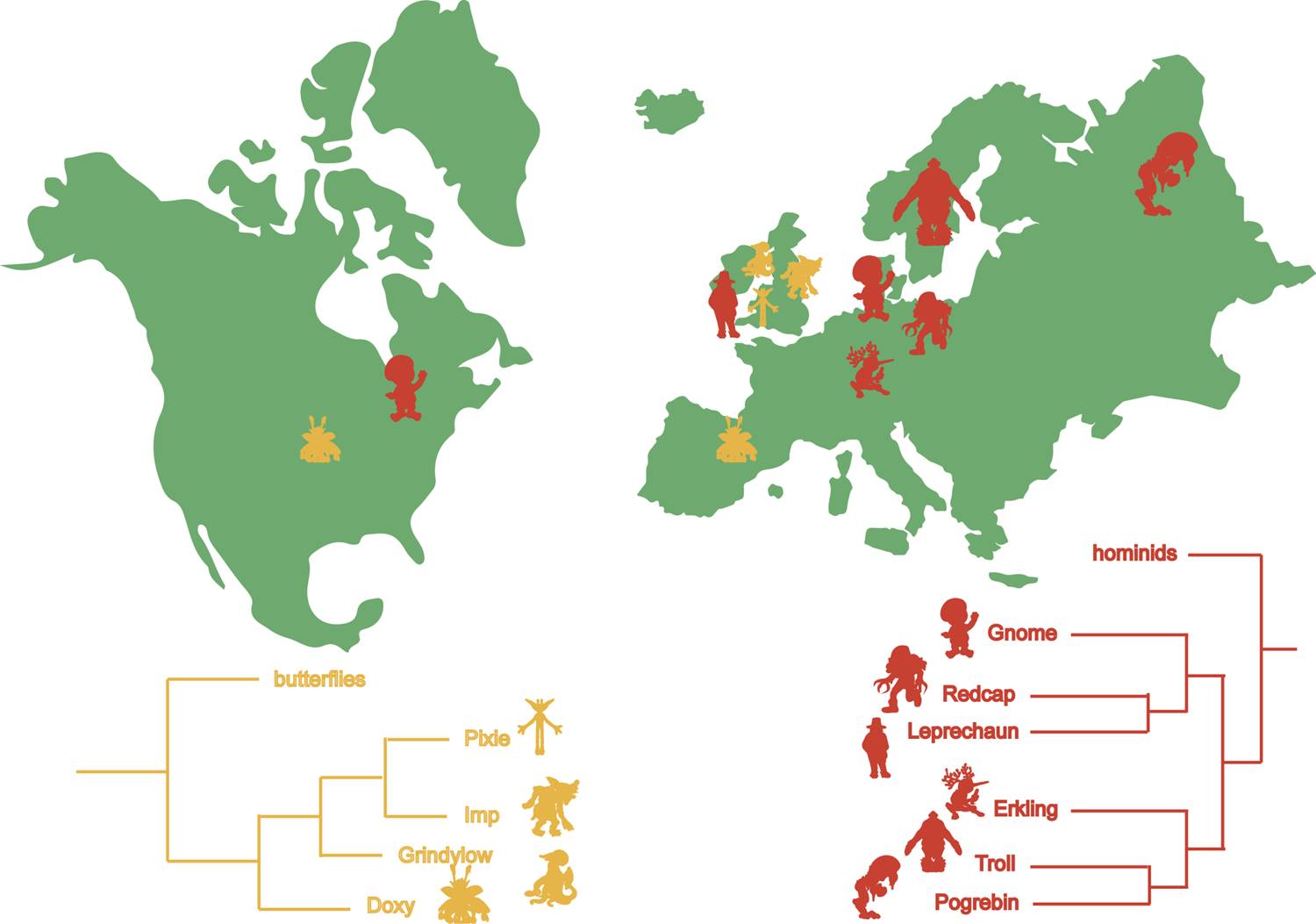
Figure 1. Current distribution of the groups discussed in the text with their phylogenetic relationships, based on Gerelle et al. (2016).
It is plausible to assume that the split between crown lepidopterans and lepidopteran-like fantastic beasts occurred back in the earliest Jurassic (Hettangian) of Britain (circa 200 Ma, i.e., 200 million years ago), as this is where the oldest fossil lepidopteran comes from (Whalley, 1986; Schachat & Gibbs, 2016). At that time, continents were united in a single land mass, called Pangaea, which would have allowed some populations of ‘Doxy-like’ beasts to migrate from British areas to what is now North America (Fig. 2A). This would explain why Doxies are present in both continents, but the remaining representatives of the group are not, demonstrating another case of disjunct distribution, as occurs, for example, with ratite birds, some pleurodiran turtles and flowering plants (Wen, 1999; de Queiroz, 2005). Otherwise, Doxies might have later migrated to North America through land continuities such as the De Geer Bridge (McKenna, 1975).
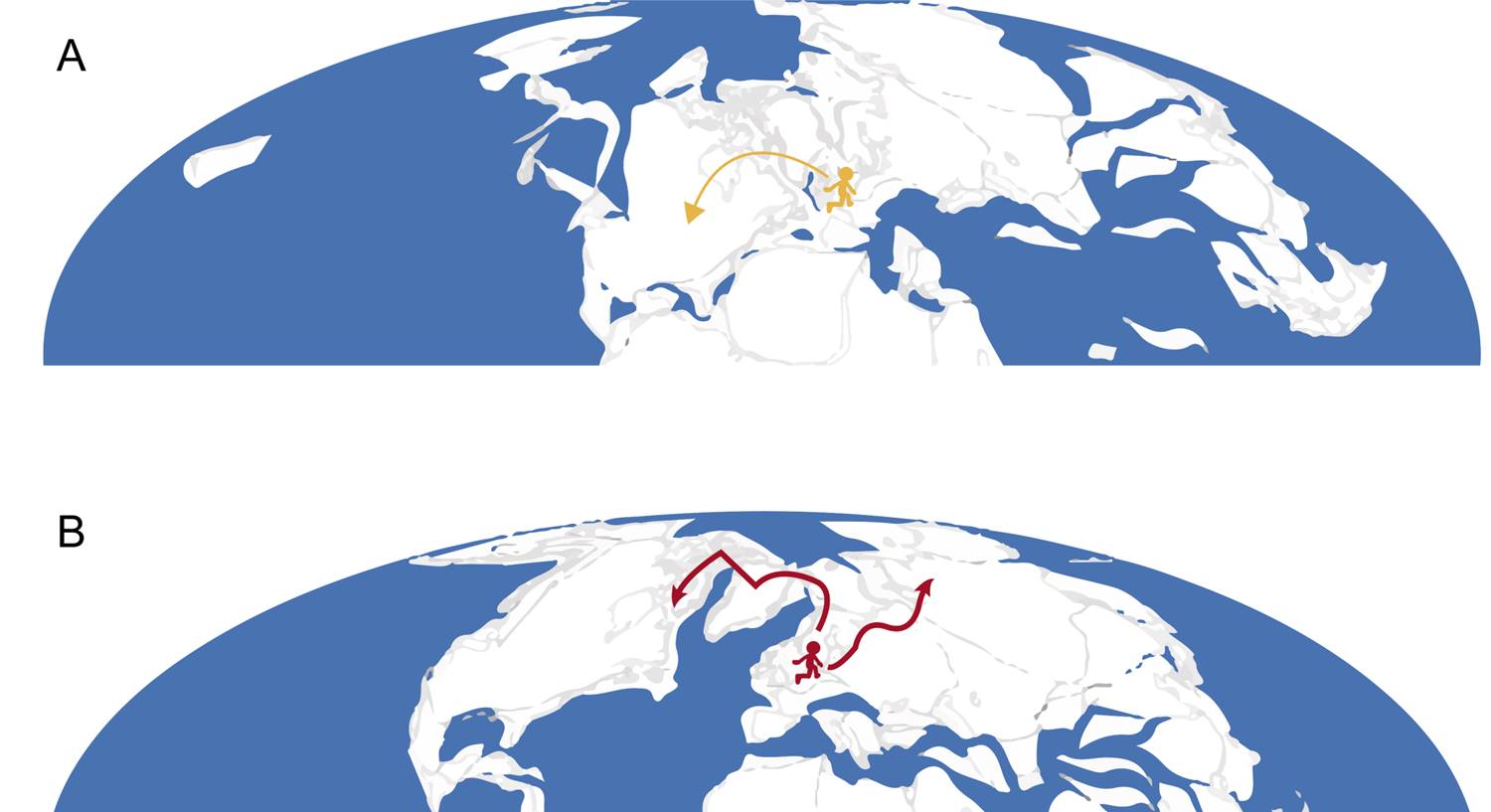
Figure 2. Probable location of ancestors of (A) the lepidopteran-like beasts during the Hettangian (earliest Jurassic) of Britain, with posterior migration to North America, and (B) hominoid-related beasts, originating in Central Europe during the Paleogene, with subsequent migration to northern Europe and North America. Maps modified from the Paleobiology Database (PBDB; www.paleobiodb.org).
Grindylows branched early in this clade’s evolutionary history, “soon” after the Doxy lineage separated, likely dating to the Toarcian (late Early Jurassic; circa 180 to 175 Ma), when England was flooded by marine transgressions (Wignall, 1991). The populations occupying the deluged area probably vanished, while the ones remaining at its borders survived and later invaded the aquatic environment (organisms closely related to modern Grindylows). This is somewhat akin to the Pleistocene refuge hypothesis of Neotropical diversification (e.g., Vanzolini & Williams, 1981; Garzón-Orduña et al., 2014), but instead of forest retraction due to climate fluctuation, areas underwent fragmentation because of marine water incursion.
Like the other splitting events, Imps and Pixies diverged mainly due to historical causes. Both beasts share morphological and reproductive similarities (Scamander, 2001). Pixies are restricted to Cornwall, whereas Imps are distributed throughout Britain, living near river banks. In Cornwall, the River Tamar largely represents the boundary with the rest of England (Carey, 1911). The rise of sea-level (similar to that of the last interglacial period; Rohling et al., 2008), could have flooded the river region, isolating populations that lived near it (like modern Imps do). On the Cornish side of the river, a small population would have differentiated, preventing gene flow after the restoration of sea levels (Fig. 3). Despite capable of flying (and thus crossing the river), Pixies are not known to form hybrids with Imps.
According to folklore, Fairies are exclusively British creatures (Briggs, 1967; Silver, 1999), but the lack of information regarding ecological preferences (Scamander, 2001), as well as fossils, hinder speculation about their evolutionary history.

Figure 3. (A) Geographical distribution of ‘Pixie + Imp’ ancestor in southwestern England. (B) Vicariant event isolating two populations and preventing gene flow. (C) Current distribution of Imps and Pixies, the latter being restricted to Cornwall.
HOMINOID-RELATED BEASTS
It is likely that, instead of historical events causing populations to split, ecological constraints were mainly responsible for the current diversity of hominoid-related beasts. The first branching lineage to be analyzed is the clade formed by Gnomes, Red Caps, and Leprechauns. As hominoid-related beasts, the group probably originated at least before the Miocene (a period spanning roughly 23 to 5 Ma; Stevens et al., 2013) and later invaded European landmasses. The burrowing habit of Gnomes most likely resulted of selective pressure due to the predation by Jarveys, a large ferret-like beast present both in Europe and North America. As such, the plesiomorphic (i.e., ancestral) condition of the group was a non-burrowing habit, which might have evolved independently in Red Caps too (Scamander, 2001). The occurrence of Gnomes in both Europe and North America depicts again a case of disjunct distribution, but the processes that drove such pattern probably differ from that of the Doxy. Rather than a vicariant event resulting from the split of Laurasia, climatological events could have created a passage that allowed them to reach North America (e.g., the Thulean Bridge; Brikiatis, 2014), as exemplified by marine diatoms during the Eocene (Bijl et al., 2013). As Jarveys intensively preyed on Gnomes, some populations likely sheltered in tunnels and acted as scavengers, feeding on the blood shed by their kin (similar to modern Red Caps).
In turn, Leprechauns likely represent a more recent lineage that migrated to Britain at first (still connected to the European mainland; Erlingsson, 2004) and then reached Ireland, probably across a land bridge before humans (Edwards & Brooks, 2008; Bower, 2016), being later included in Irish folklore (Winberry, 1976; Koch, 2006). However, Leprechauns (as all the exemplified beasts) lack a fossil record, which complicates the understanding of how and when such groups colonized the areas they currently live in (Crottini et al., 2012).
The other clade of hominoid-related beasts comprises Erklings, Trolls and Progebins, distributed in northern Europe (Fig. 4A). Modern representatives of the group are known to feed on flesh (especially human; Scamander, 2001), which evokes whether such beasts arose earlier or later than the Homo arrival to Europe (ca. 1.4–1.8 Ma; Parfitt et al., 2005; Toro-Moyano et al., 2013). Probably spread all over Europe originally, the competition for the same kind of resources (mostly raw flesh) with a distantly related clade

Figure 4. (A) Probable ancient distribution of Erklings, Trolls, and Pogrebins in Europe. (B) Arrival of Homo species in Europe, ca. 1.5 Ma. (C) Demise of original populations of fantastic creatures, showing their current relictual distribution in Europe.
(Homo species) may have constrained the range of the group (mainly inhabiting densely vegetated zones today), extinguishing ancient populations more widely distributed. This last example analogously illustrates a case (e.g., Silvestro et al., 2015) in which the later arrival of a phylogenetically distant (but ecologically similar) clade to an area triggered diversification shifts onto the previous occupiers, as well as the probable extinction of some forms.
BIOGEOGRAPHICAL HISTORY OF FANTASTIC BEASTS
In order to verify if there is a regionalization among the fantastic biota, their geographical distribution was compiled from Scamander (2001) and interpreted based on (i) six distinct geographical realms from Wallace (1876), and (ii) the recent division of Holt et al. (2013) in 13 domains. Each creature was plotted against the realm in a simple area vs. taxa matrix (e.g., Souza, 2005), scoring (0) if absent, and (1) if present in a determined locality. This gives us a diagram, called ‘area cladogram’, with the biogeographic history of the groups.
The area cladogram obtained with Wallace’s six biogeographic domains (Wallace, 1876) is partially consistent with the biogeographical history of the southern hemisphere (i.e., mostly Gondwanan-derived land masses), according to patterns observed in some plants and animals (e.g., Sanmartín & Ronquist, 2004), in which the Oriental biota (i.e., mainly Indian) is the sister group to the remaining areas (Fig. 5A). This could be reasonably expected, since India was the first land mass to branch in Gondwana breakup geological sequence (Barron, 1987; McLoughlin, 2001). The relationships of African, South American and Australian areas however disagree with Sanmartín & Ronquist (2004), in which it was expected that South American and Australian biotas were more closely related to one another than to the African biota. This result could imply a Pangaean origin for these fantastic beasts, with subsequent vicariant events. However, this hierarchical pattern following the breakup sequence of Gondwana could also be a kind of ‘vicariance-mimicking’ phenomenon affecting the cladogram area topology (see Upchurch et al., 2002). Until fossils of fantastic beasts are found, knowledge about their past distribution remains obscured. On the other hand, when plotted according to the biogeographic realms of Holt et al. (2013) the Gondwanan-derived continents do not present such hierarchical relationship (Fig. 5B), resulting in a pectinate (i.e., comb-like) conformation within the area cladogram. Both results could also be influenced by the lack of data about the fantastic beasts, which may not follow the pattern of ordinary ones.
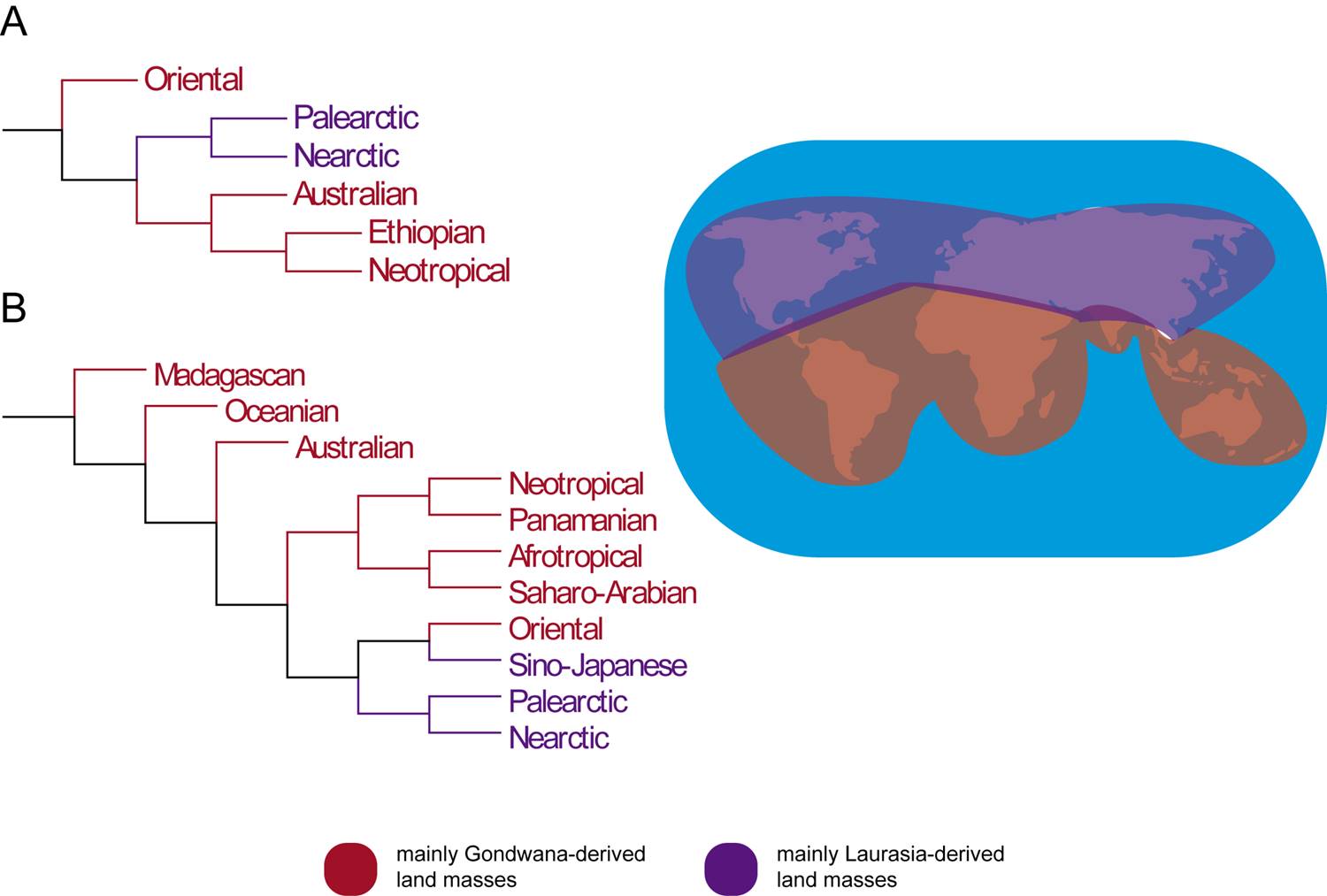
Figure 5. Area cladograms obtained based on (A) Wallace’s zones (1876), and (B) Holt et al. (2013) new zones, subdividing those proposed by Wallace.
In sum, due to the incompatible results for Gondwanan continents, the fantastic biota could have had a hybrid, composed origin (Amorim, 2012), with both autochthonous and allochthonous elements. The Palearctic and Nearctic realms were recovered together in both analyses, although both regions are inhabited by most of the beasts, which could have biased the result. Despite of the apparently unarguable Laurasian distribution of such beasts, it has been historically difficult to depict the continents’ biogeographical scenario (Sanmartín et al., 2001; Wildman et al., 2007).
FICTION AS SCIENCE OUTREACH
Biogeography is an integrative science combining different sources of evidence to understand what caused organisms to be distributed the way they presently are – or were in the geological past (Lomolino et al., 2010). Despite of its relevance, the public knowledge (i.e., outside the academic environment) concerning this research area seems debilitated, even with the timid increase in electronic dissemination (Ladle, 2008). Present in both evolutionary approaches of Darwin (1859) and Wallace (1876), the spatial distribution of organisms offers an unparalleled tool to stimulate students to think about evolution and natural history (Rosenau, 2012; Allchin, 2014) – and not just to understand evolution, but to accept it as well (Lombrozo et al., 2008).
In this context, the teaching of biogeography (and evolution in general) could benefit from the use of fictional organisms with “real” distributions around the globe. Presenting the continents’ past and present arrangement, allied with the localities inhabited by the beasts and possible disjunction events, in a kind of inquiry-based approach (e.g., Robbins & Roy, 2007) would instigate students to formulate their own hypotheses. This, in turn, could lead them to more easily assimilate all these concepts. The specific use of the popular Fantastic Beasts of the Harry Potter franchise to canalize this is supoprted mostly by the interest of younger audiences (under 25 years old) in the recently released spin-off movie (over 50%; Lang, 2016). Actually, scientific scenarios were already present on several episodes from the Harry Potter books (e.g., Rowling, 1997; 1998; 1999; 2005), providing a larger background for people to get involved.
Moreover, this would not be the first time that a fictional universe was considered to engage younger people on scientific activities (e.g., Roque, 2016). J.K. Rownling’s fantasy novels are already proven as a promising and innovative background for scientific experiments (e.g., Vezzali et al., 2014). As such, the present work is hopefully in a good position to arouse at least a spark of interest among students to understand what made our beasts – fantastic or otherwise – to live where they do.
REFERENCES
Allchin, D. (2014) On genius and happenstance in scientific discovery. The American Biology Teacher 76: 145–148.
Alroy, J.; Koch, P.L.; Zachos, J.C. (2000) Global climate change and North American mammalian evolution. Paleobiology 26: 259–288.
Amorim, D.S. (2012) Biogeografia da Região Neotropical. In: Rafael, J.A.; Melo, G.A.R.; Carvalho, C.J.B.; Casari, S.A.; Constantino, R. (Eds.) Insetos do Brasil: Diversidade e Taxonomia. Editora Holos, Ribeirão Preto. Pp. 111–132.
Barron, E.J. (1987) Cretaceous plate tectonics reconstructions. Palaeogeography, Palaeoclima-tology, Palaeoecology 59: 3–29.
Bijl, P.K.; Bendle, J.A.; Bohaty, S.M.; Pross, J.; Schouten, S.; Tauxe, L.; Stickley, C.E.; McKay, R.M.; Röhl, U.; Olney, M.; Sluijs, A.; Escutia, C.; Brinkhuis, H.; Expedition 318 Scientists. (2013) Eocene cooling linked to early flow across the Tasmanian Gateway. PNAS 110: 9645–9650.
Briggs, K.M. (1967) The Fairies in English Tradition and Literature. University of Chicago Press, Chicago.
Brikiatis, L. (2014) The De Geer, Thulean and Beringia routes: key concepts for understanding early Cenozoic biogeography. Journal of Biogeography 41: 1036–1054.
Bower, B. (2016) Bear bone rewrites human history in Ireland. Available from: https://www. sciencenews.org/article/bear-bone-rewrites-hu man-history-ireland (Date of access: 02/Nov/ 2016).
Carey, W.M. (1911) The geography of Cornwall. The Geographical Teacher 6: 90–103.
Crottini, A.; Madsen, O.; Poux, C.; Strauß, A.; Vieites, D.R.; Vences, M. (2012) Vertebrate time-tree elucidates the biogeographic pattern of a major biotic change around the K-T boundary in Madagascar. PNAS 109: 5358–5363.
Darwin, C.R. (1859) On the Origin of Species by Means of Natural Selection. John Murray, London.
de Queiroz, A. (2005) The resurrection of oceanic dispersal in historical biogeography. Trends in Ecology and Evolution 20: 68–73.
d’Huy, J. (2013) Le motif du dragon serait paléolithique: mythologie et archéologie. Préhistoire du Sud-Ouest 21(2): 195–215.
Edwards, R.J & Brooks, A.J. (2008) The island of Ireland: drowning the myth of an Irish land-bridge? In: Davenport, J.J.; Sleeman, D.P.; Woodman, P.C. (Eds.) Mind the Gap: Postglacial Colonisation of Ireland. Special Supplement to the Irish Naturalists’ Journal, Dublin. Pp. 19–34.
Erlingsson, U. (2004) Atlantis from a Geographer’s Perspective. Lindorm Publishing, Miami.
Garzón-Orduña, I.J.; Benetti-Longhini, J.E.; Brower, A.V.Z. (2014) Timing the diversification of the Amazonian biota: butterfly divergences are consistent with Pleistocene refugia. Journal of Biogeography 41: 1631–1638.
Gerelle, W.; Scamander, N.; Vahanvaty, A. (2016) Preliminary phylogeny of magical and ordinary creatures: evidence of a recent diversification. Available from: http://www.scq.ubc.ca/wp-con tent/uploads/2015/03/APCMvol2paper01_HarryPotter_Wesley_Ammar.pdf (Date of access: 01/Nov/2016).
Hamilton, A.J.; May, R.R.; Waters, E.K. (2015) Zoology: here be dragons. Nature 520: 42–43.
Holt, B.G.; Lessard, J.-P.; Borregaard, M.K.; Fritz, S.A.; Araújo, M.B.; Dimitrov, D.; Fabre, P.-H.; Graham, C.H.; Graves, G.R.; Jønsson, K.A.; Nogués-Bravo, D.; Wang, Z.; Whittaker, R.J.; Fjeldså, J.; Rahbe, C. (2013) An update of Wallace’s zoogeographic regions of the world. Science 339: 74–78.
Janis, C.M. (1993) Tertiary mammal evolution in the context of changing climates, vegetation, and tectonic events. Annual Review of Ecology and Systematics 24: 467–500.
Koch, J.T. (2006) Celtic Culture: a Historical Encyclopedia. ABC-CLIO, Santa Barbara.
Kukalová-Peck, J. (1991) Fossil history and the evolution of the hexapod structures. In: Naumann, I.D. (Ed.) The Insects of Australia: a Textbook for Students and Research Workers. Melbourne University Press, Melbourne. Pp. 141–179.
Ladle, R.J. (2008) Catching fairies and the public representation of biogeography. Journal of Biogeography 35: 388–391.
Lang, B. (2016) ‘Fantastic Beasts’ box office debut draws on aging ‘Harry Potter’ fanbase. http://variety.com/2016/film/box-office/fantas tic-beasts-box-office-harry-potter-1201923148/ (Date of access: 20/Nov/2016).
Lombrozo, T.; Thanukos, A.; Weisberg, M. (2008) The importance of understanding the nature of science for accepting evolution. Evolution: Education and Outreach 1: 290–298.
Lomolino, M.V.; Riddle, B.R.; Whittaker, R.J.; Brown, J.H. (2010) Biogeography. Sinauer Associates, Sunderland.
McKenna, M.C. (1975) Fossil mammals and early Eocene North Atlantic land continuity. Annals of the Missouri Botanical Garden 62: 335–353.
McLoughlin, S. (2001) The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism. Australian Journal of Botany 49: 271–300.
Parfitt, S.A.; Barendregt, R.W.; Breda, M.; Candy, I.; Collins, M.J.; Coope, G.R.; Durbidge, P.; Field, M.H.; Lee, J.R.; Lister, A.M.; Mutch, R.; Penkman, K.E.H.; Preece, R.C.; Rose, J.; Stringer, C.B.; Symmons, R.; Whittaker, J.E.; Wymer, J.J.; Stuart, A.J. (2005). The earliest record of human activity in northern Europe. Nature 438: 1008–1012.
Robbins, J.R. & Roy, P. (2007) The natural selection: identifying and correcting non-science student preconceptions through an inquiry-based, critical approach to evolution. The American Biology Teacher 69: 460–466.
Rohling, E.J.; Grant, K.; Hemleben, C.; Siddall, M.; Hoogakker, B.A.A.; Bolshaw, M.; Kucera, M. (2008) High rates of sea-level rise during the last interglacial period. Nature Geoscience 1: 38–42.
Roque, F.O. (2016) Field studies: could Pokemon Go boost birding? Nature 537: 34–34.
Rosenau, J. (2012) Evolution and biogeography: leading students in Darwin and Wallace’s footsteps. Evolution: Education and Outreach 5: 582–584.
Rowling, J.K. (1997) Harry Potter and the Philosopher’s Stone. Bloomsbury, London.
Rowling, J.K. (1998) Harry Potter and the Chamber of Secrets. Bloomsbury, London.
Rowling, J.K. (1999) Harry Potter and the Prisoner of Azkaban. Bloomsbury, London.
Rowling, J.K. (2005) Harry Potter and the Half-Blood Prince. Bloomsbury, London.
Sanmartín, I.; Enghoff, H.; Ronquist, F. (2001) Patterns of animal dispersal, vicariance and diversification in the Holarctic. Biological Journal of the Linnean Society 73: 345–90.
Sanmartín, I.; Ronquist, F. (2004) Southern hemisphere biogeography inferred by event-based models: plant versus animal patterns. Systematic Biology 53: 216–243.
Scamander, N. (2001) Fantastic Beasts and Where to Find Them. Bloomsbury, London.
Schachat, S.R & Gibbs, G.W. (2016) Variable wing venation in Agathiphaga (Lepidoptera: Agathiphagidae) is key to understanding the evolution of basal moths. Royal Society Open Science 3: 160453.
Silver, C.G. (1999) Strange and Secret Peoples: Fairies and Victorian Consciousness. Oxford University Press, Oxford.
Silvestro, D.; Antonelli, A.; Salamin, N.; Quental, T.B. (2015) The role of clade competition in the diversification of North American canids. PNAS 112: 8684–8689.
Souza, F.L. (2005) Geographical distribution patterns of South American side-necked turtles (Chelidae), with emphasis on Brazilian species. Revista Española de Herpetología 19: 33–46.
Stevens, N.J.; Seiffert, E.R.; O’Connor, P.M.; Roberts, E.M.; Schmitz, M.D.; Krause, C.; Gorscak, E.; Ngasala, S.; Hieroymus, T.L.; Temu, J. (2013) Palaeontological evidence for an Oligocene divergence between Old World monkeys and apes. Nature 497: 611–614.
Toro-Moyano, I.; Martínez-Navarro, B.; Augustí, J.; Souday, C.; Castro, J.M.B.; Martinón-Torres, M.; Fajardo, B.; Duval, M.; Falguères, C.; Oms, O.; Parés, J.M.; Anadón, P.; Julià, R.; García-Aguilar, J.M.; Moigne, A.-M.; Espigares, M.P.; Ros-Montoya, S.; Palmqvist, P. (2013) The oldest human fossil in Europe dated to ca. 1.4 Ma at Orce (Spain). Journal of Human Evolution 65: 1–9.
Upchurch, P.; Hunn, C.A.; Norman, D.B. (2002). An analysis of dinosaurian biogeography: evidence for the existence of vicariance and dispersal patterns caused by geological events. Proceedings of the Royal Society B: Biological Sciences 269: 613–621.
Vanzolini, P.E. & Williams, E.E. (1981) The vanishing refuges: a mechanism for ecogeographic speciation. Papéis Avulsos de Zoologia 34: 251–255.
Vezzali, L.; Stathi, S.; Giovannini, D.; Capozza, D.; Trifiletti, E. (2014) The greatest magic of Harry Potter: reducing prejudice. Journal of Applied Social Psychology 45: 105–121.
Wallace, A.R. (1876) The Geographical Distribution of Animals. Cambridge University Press, Cambridge.
Wen, J. (1999) Evolution of eastern Asian and eastern North American disjunct distribution in flowering plants. Annual Review of Ecology and Systematics 30: 421–455.
Whalley, P. (1986) A review of the current fossil evidence of Lepidoptera in the Mesozoic. Biological Journal of the Linnean Society of London 28: 253–271.
Wignall, P.B. (1991) Model for transgressive black shales? Geology 19: 167–170.
Wildman, D.E.; Uddin, M.; Opazo, J.C.; Liu, G.; Lefort, V.; Guindon, S.; Gascuel, O.; Grossman, L.I.; Romero, R.; Goodman, M. (2007) Genomics, biogeography, and the diversification of placental mammals. PNAS 104: 14395–14400.
Winberry, J.J. (1976) The elusive elf: some thoughts on the nature and origin of the Irish leprechaun. Folklore 87: 63–75.
ACKNOWLEDGMENTS
I would like to thank J.K. Rowling, who idealized the magical world of Harry Potter as well as its fantastic creatures; my Biogeography professor, Eduardo Almeida, in whose course I was able to formulate ideas regarding the subject; my colleagues (Anaís Silveira, Carolina Barroso, Fernanda Dalarmi, Isabela Soares, Luene Pessoa e Thayná Medeiros) with whom I worked in said course, as well as Nádia Gibran, for all the support and kindness. The author is funded by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP; proc. 2016/03373-0).
ABOUT THE AUTHOR
Guilherme Hermanson is a big fan of the Harry Potter magical world. He is also an undergraduate student at the University of São Paulo (Ribeirão Preto’s campus), currently developing his research at the university’s Paleontology Lab, focused on the internal anatomy of extinct turtles.