Skip to content
RSS Feed
Bluesky
Type your email…
Subscribe
Search
Journal of Geek Studies
About
Guidelines for authors
Current Issue
Archives
Vol. 13(1): Jun/2026
Vol. 12(2): Dec/2025
Vol. 12(1): Jun/2025
Vol. 11(2): Dec/2024
Vol. 11(1): Jun/2024
Older issues
Editorial Team
Partners
Contact
July 19, 2026
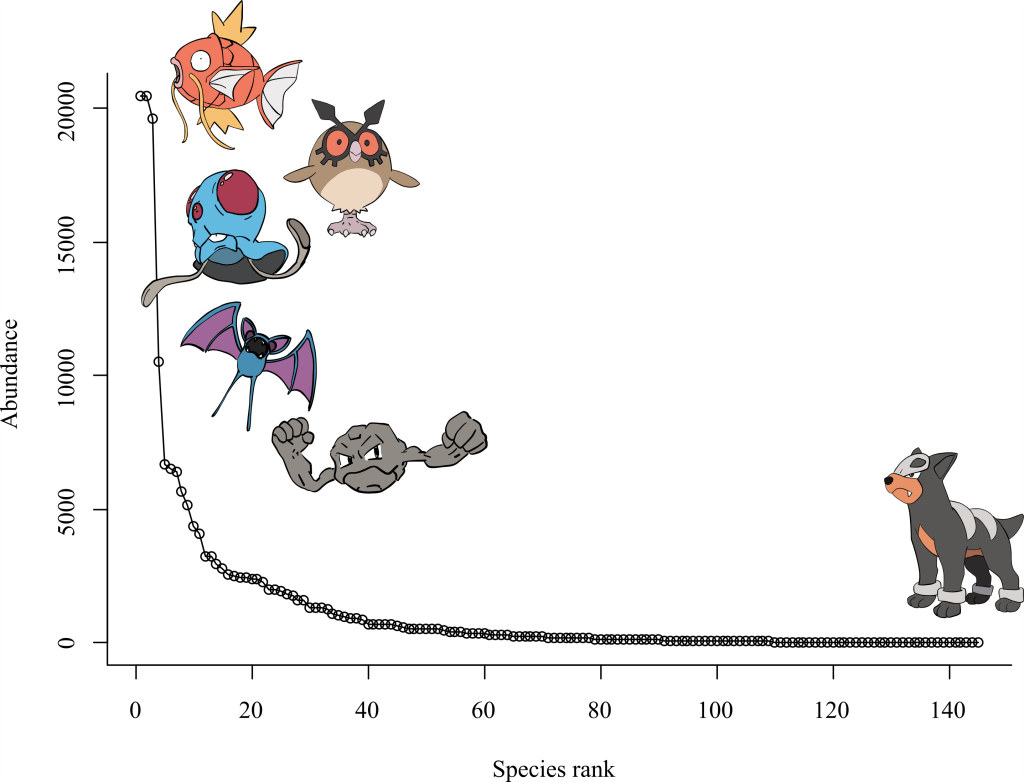
Ecological patterns in Pokémon games reflect those observed in the natural world
Biology
,
Video Games
Ecology
,
Pokémon
,
Zoology
April 16, 2026
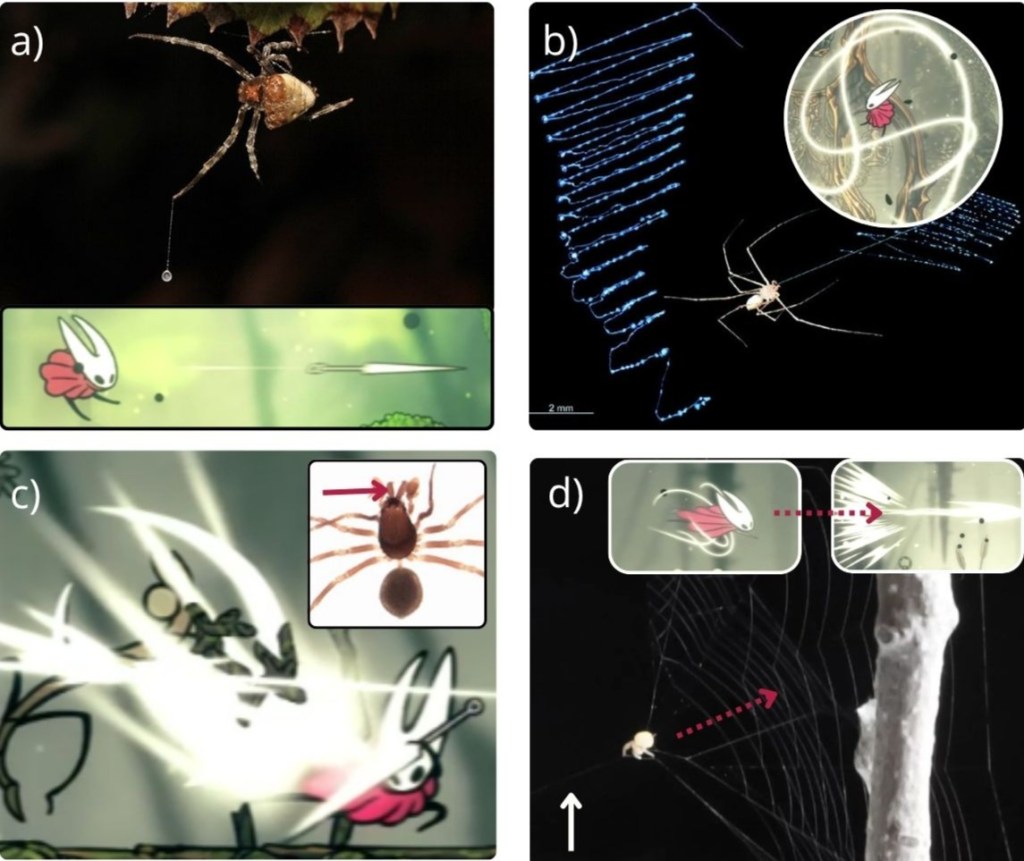
Hornet’s web: weaving the biology of silk into Hollow Knight: Silksong
Biology
,
Video Games
,
Zoology
Arachnology
,
Behaviour
,
Ecology
,
Hollow Knight
,
Metroidvania
February 5, 2026
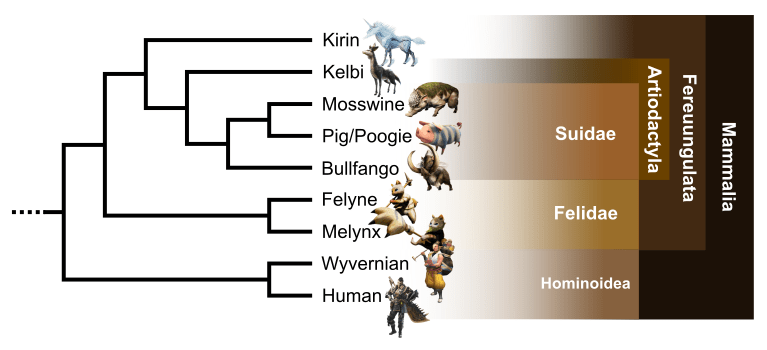
Monster Hunter: an evolutionary story
Biology
,
Video Games
,
Zoology
Monster Hunter
,
Taxonomy
,
Zoology
January 17, 2026
Analysis of Eirin Yagokoro (Touhou Project) from a bioethical perspective
Jurisprudence
,
Medicine
,
Video Games
Ethics
,
Touhou
November 11, 2025
Can conservation really be effective through video games? A case study on Jack Barau and some perspectives
Biology
,
Education
,
Environmental Sciences
,
Video Games
,
Zoology
Ecology
,
Game Development
,
Nature Conservation
,
Ornithology
,
Zoology
November 1, 2025
When trash stops being a treasure: pollution and its effects in Another Crab’s Treasure
Biology
,
Environmental Sciences
,
Video Games
Carcinology
,
Ecology
,
Marine Biology
,
Nature Conservation
,
Zoology
September 19, 2025
When cyberpunk leaks into the timeline: megacorps, memes, and Rheinmetall as a real‑world case study
Cinema
,
Science Fiction
,
Sociology
,
Video Games
Cyberpunk
,
Memetic Discourse
,
Political Science
,
Science Fiction
July 11, 2025
Predicting competitive Pokémon VGC leads using Latent Semantic Analysis: a data-driven approach to team matchups
Artificial Inteligence
,
Simulation
,
Video Games
Natural language processing
,
Pokémon
May 1, 2025
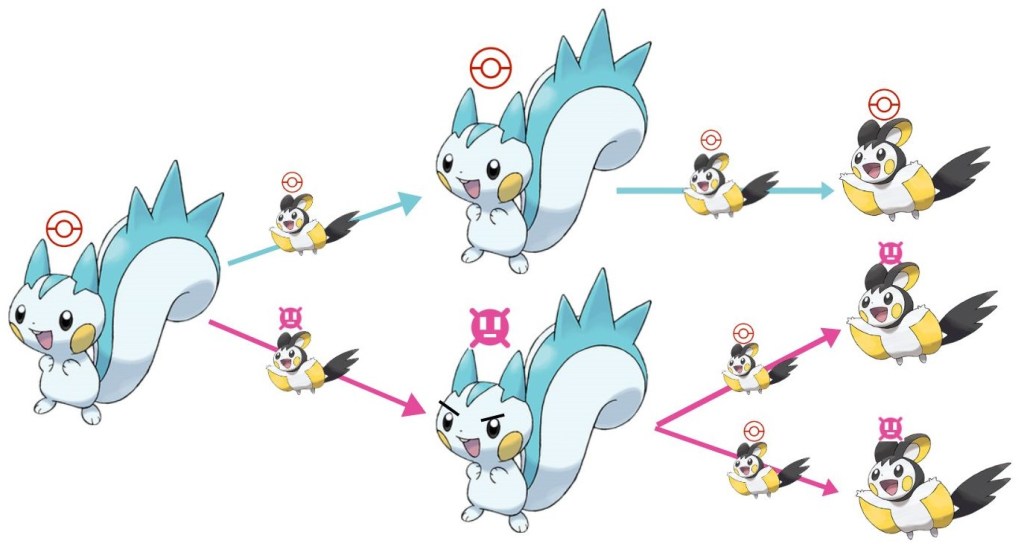
Pokérus: unraveling the biology of the Pokémon virus
Biology
,
Video Games
Pokémon
,
Virology
April 5, 2025
Archaeolinguistics and language deciphering in action-adventure video games: Indiana Jones, Tomb Raider and Uncharted
Archaeology
,
Cinema
,
Video Games
Archaeolinguistics
,
Archaeology
,
Indiana Jones
,
Tomb Raider
,
Uncharted
1
2
3
…
9
Next Page
Subscribe
Subscribed
Journal of Geek Studies
Join 309 other subscribers
Sign me up
Already have a WordPress.com account?
Log in now.
Journal of Geek Studies
Subscribe
Subscribed
Sign up
Log in
Report this content
View site in Reader
Manage subscriptions
Collapse this bar