Skip to content
RSS Feed
Bluesky
Type your email…
Subscribe
Search
Journal of Geek Studies
About
Guidelines for authors
Current Issue
Archives
Vol. 13(1): Jun/2026
Vol. 12(2): Dec/2025
Vol. 12(1): Jun/2025
Vol. 11(2): Dec/2024
Vol. 11(1): Jun/2024
Older issues
Editorial Team
Partners
Contact
Fantastic Beasts
November 22, 2025
Steampunk dinosaurs and Victorian-era palaeontology
Anime & Manga
,
Biology
,
History
Dinosaurs
,
Fantastic Beasts
,
Paleontology
,
Steampunk
March 8, 2025
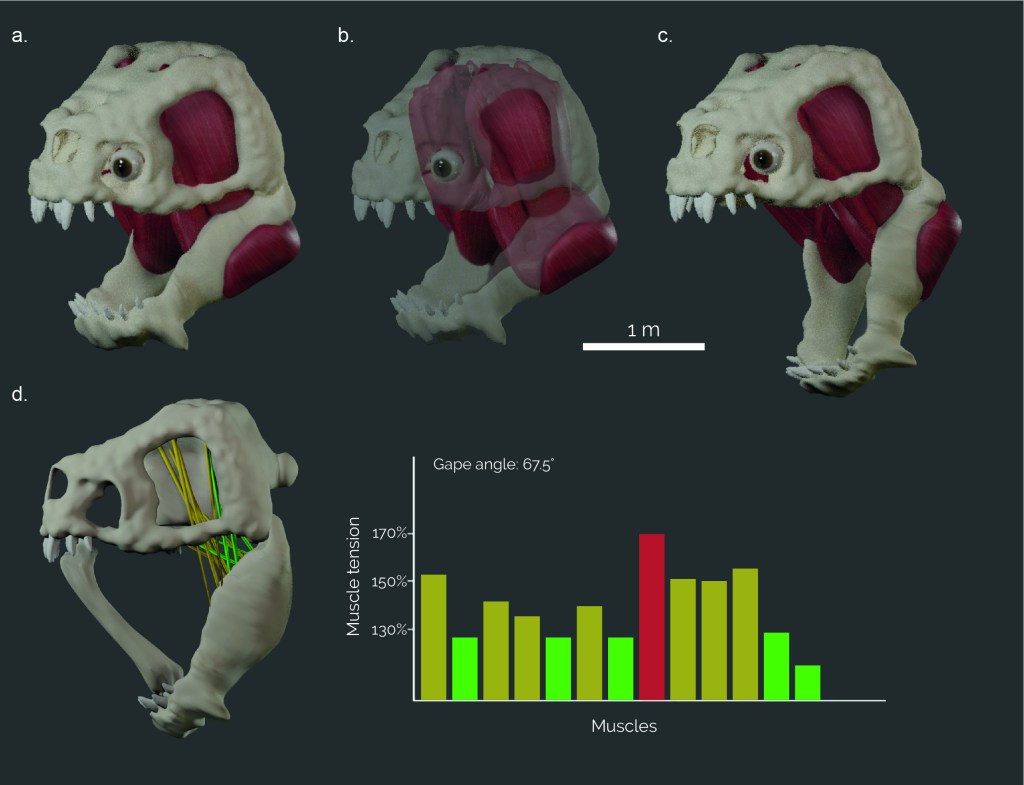
Is a bone a viable weapon when combating a Rancor? Estimating the bite force of an intergalactic mega-predator
Biology
,
Cinema
,
Paleontology
,
Physics
,
Television Series
Dynamics
,
Fantastic Beasts
,
Finite element method
,
Science Fiction
,
Star Wars
December 2, 2024
Xenomorphia ex machina: the zoology and biochemistry of xenomorphs from the Alien franchise
Biology
,
Cinema
,
Horror
,
Zoology
Alien
,
Fantastic Beasts
,
Horror Movies / Books
,
Science Fiction
,
Taxonomy
,
Xenomorphs
November 11, 2023
My neighbor Linnaeus: The influence of Studio Ghibli in zoological nomenclature
Anime & Manga
,
Biology
,
Cinema
,
Japan Studies
,
Zoology
Ecology
,
Fantastic Beasts
,
Ghibli
,
Taxonomy
,
Zoology
September 17, 2023
The magic of transformation: a comparative analysis of alchemy in Harry Potter and its real-world chemistry roots
Chemistry
,
Cinema
,
Literature
,
Mythology
Alchemy
,
Chemistry
,
Fantastic Beasts
,
Harry Potter
August 15, 2023
Unleashing the Alien: a deep dive into the terrifying world of Xenomorphs in science fiction and horror
Cinema
,
Horror
Alien
,
Fantastic Beasts
,
Horror Movies / Books
,
Science Fiction
,
Xenomorphs
August 5, 2021
Short note on recent sightings of Rodan in Godzilla Singular Point
Anime & Manga
,
Biology
,
Cinema
,
Paleontology
Ecology
,
Fantastic Beasts
,
Giant Monsters
,
Godzilla
,
Taxonomy
,
Zoology
February 23, 2021
Using How to Train Your Dragon to teach about endangered species, the scientific method, and popularization of science
Animation
,
Biology
,
Cinema
Dragons
,
DreamWorks
,
Ecology
,
Fantastic Beasts
,
Zoology
August 26, 2018
Zoological Nomenclature of Ice and Fire
Biology
,
Literature
,
Television Series
Fantastic Beasts
,
Game of Thrones
,
Zoological Nomenclature
,
Zoology
September 3, 2017
Making a vampire
Biology
,
Cinema
,
Horror
,
Literature
,
Mythology
Fantastic Beasts
,
Genetics
,
Horror Movies / Books
,
Vampires
1
2
Next Page
Subscribe
Subscribed
Journal of Geek Studies
Join 309 other subscribers
Sign me up
Already have a WordPress.com account?
Log in now.
Journal of Geek Studies
Subscribe
Subscribed
Sign up
Log in
Report this content
View site in Reader
Manage subscriptions
Collapse this bar