Andrea Villa1 & Alessio del Negro2
1Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Barcelona, Spain. Email: andrewjayhouse (at) gmail (dot) com
2Independent Researcher, Milano, Italy. Email: alessio.delnegro05 (at) gmail (dot) com
https://doi.org/10.5281/zenodo.8226258
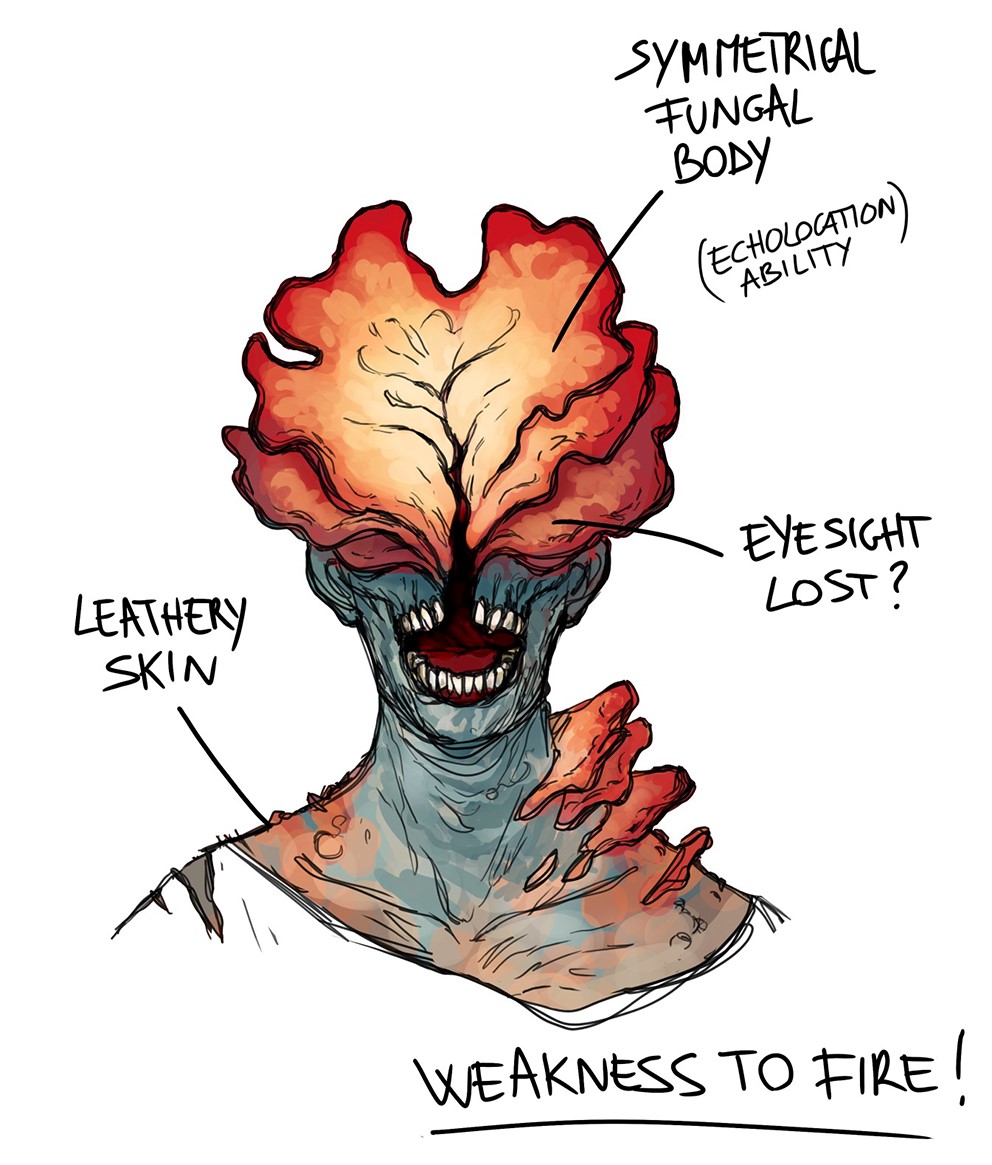
Starting in 2013 and recently back to hit the headlines with the sequel, The Last of Us (TLoU, herein referring to either the franchise as a whole or the first game in the series) is one of the leading franchises created by the software house Naughty Dog. This award-winning series of horror-themed action-adventure games takes place in a dystopic future where civilization collapsed due to an emergent disease caused by a parasitoid fungus that transforms humans into violent zombie- or better ghoul-like creatures (Fig. 1).
The disease was called Cordyceps Brain Infection (CBI), and is reported as originating from a new species of the genus Cordyceps that evolved to infect humans instead of arthropods, as usual for these parasitoid fungi (see medical pamphlet published by the Center of Disease Control found in the outskirts of Boston in TLoU). While some information on the CBI were already given within the games, that was mainly resulting from studying the disease and its effects under a medical point of view. Less attention was given to the fungal infection under its biological and evolutionary aspects (somehow understandably given that most of the knowledge was gathered under emergency conditions and trying to oppose a pandemic of unprecedented violence).
We explore here the CBI-infected using a natural science approach, with special focus on their morphological and especially ethological or behavioural features. Our main aim is to shed some light on the possible functional and evolutionary significance of those features.
METHODOLOGY
The main source of the morphological data presented herein are the 3D models of the CBI infected available as additional content in The Last of Us Part II (TLoU2). Information of the ethology of the infected was observational data retrieved from encounters in both TLoU and TLoU2. The Last of Us Wiki on Fandom (https://thelastofus.fandom.com/it/wiki/The_Last_of_Us_Wiki) was used as a supplementary source of information.
In this article, we take the stance of researchers analysing the infected as if we were in the reality of TLoU, taking the game’s world at face value. We draw parallels with real-world species and phenomena to enrich our explanations and explore the possibilities of that fictional world.
RESULTS
Six different types of infected are currently known. They represent successive stages of the infection, except for shamblers that might be an alternative stage to bloaters (possibly related to a more humid environment; see the Shambler Note in TLoU2, but also Turi, 2019) and the Rat King, whose real nature is still not clear. The fungus growth seems to give increased strength and physical resistance to the host with the progression of the infection.
Runners
Runners (Fig. 2A) are the simplest kind of infected, representing the first stage of the CBI. They still have a generally human aspect, with few appreciable changes. Lesions are usually present on the body, but it is not always clear whether these may be due to the fungus or just the accidental results of a lower self-care of the host. Hyphae are visible through the skin. Very dark blood (or another kind of fluid) pours out from head orifices. The eyes are strongly reddened and the dermis has a more or less vaguely ashy colour. They are still capable of both hearing and seeing. Behavioural features include relatively fluid movements, vocalizations (mainly screams and moans), and capacity of walking large distances. They form hordes and are found in both indoor and outdoor environments.
Stalkers
The looks of these infected is still vaguely human (Fig. 2B), but with a more decomposed appearance: e.g., more lacerated skin, unhealthier colour. This is likely due to the longer time elapsed since the infection. Hyphae are starting to come out from the host body. The exposed hyphae are mainly located in the upper part of the body: the head, where the most mature fungi are, and also the shoulders. Leathery areas of the skin start to appear in sparse parts of the body. The behaviour includes relatively fluid movements and hiding/ambush habits. They still react to both auditory and visual stimuli, and are also capable of rudimentary echolocation. They are usually encountered in small groups, but it is not known if they may also form hordes. They are mostly found indoors, but sometimes out in the open as well, in particular at night or in darker areas.
Clickers
Clickers (Figs. 1, 2C) are still human-like in body shape, but their skin is now completely leathery. Real hyphae are not visible anymore on the body, but the body is overall bulging. Like the hyphae in other infected, these bulges are also more concentrated in the upper part of the trunk. The skull is broken by a large fungal body coming out to cover the entire upper part of the face. This fungal body has an almost symmetrical shape, with two lobes flanking a sort of median vertical opening through which the clicker takes advantage of its echolocation abilities. The broken head and the fungal body covering the face prevent clickers from seeing, even though functional eyes may still be present in at least some of them (as it seems to be testified by their reaction to visual stimuli when the fungal body is damaged or artificially removed). In any case, they react to auditory stimuli and are able to echolocate. Their movements are less fluid than that of runners and stalkers, with twitchy movements particularly while walking. They can be found in small groups, both indoors and outdoors, but they also aggregate in hordes.
Shamblers
Shamblers (Fig. 2D) have a human-like aspect, but a leathery skin and a swollen physique. They are larger than other infected, but not as large as bloaters. Subcutaneous hyphae are vaguely visible here and there, especially on the head. Fungal bodies similar to arboreal fungi are present on the arms, legs, and part of the trunk, distributed principally on the posterolateral sides. Rounded pomegranate-like structures filled with corrosive ichor and gas accumulate between the shoulders and the head, without covering the face. Shamblers can detach and throw these rounded structures, which explode upon contact, releasing their content. The facial area is usually broken or damaged, but not in the same way as the clickers (i.e., there is no emerging fungal body). Eyes are still present and functional. Shamblers are slow, but they do not show the same twitchy moves as clickers do. They can chase efficiently with rather rapid runs. These infected are apparently restricted to small delimited areas, and it seems that they do not gather in hordes. They are usually encountered in very small groups of one or two individuals, together with other kinds of infected.
Bloaters
These infected (Fig. 2E) are larger than shamblers. They have a swollen humanoid appearance, with their skin completely covered by different kinds of fungal structures: filiform hyphae, rounded structures (similar to those of the shamblers, but located instead on their limbs and trunk), bulges, and more elongated structures. The skull is broken and almost completely obscured by an emerging fungal body, as in clickers. There are no visible eyes anymore, thus suggesting hearing/echolocation-mediated sensorial activity. Echolocation in bloaters is indeed well developed. Like shamblers, they can throw the rounded structures present on their body. They are generally found alone or with few infected of other kinds, in closed places. They do not form hordes.

Rat King
This is a very peculiar infected (Fig. 3), currently only known from a single individual in the depths of the hospital in Seattle (the quarantine area). Because of this, it is possible that this is not a real stage of the infection, but rather the outcome of a random event that was made possible by the conditions present in that location (i.e., numerous infected stored together in the same closed space for a long amount of time). The Rat King is very large, with an overall deformed humanoid aspect. It apparently results from the merging of numerous infected from which it takes, without a real regular pattern, the main features. Its components include a bloater, at least one stalker (Fig. 4), and various clickers. The bloater and the stalker, at least, are still able to act independently.


DISCUSSION
The modified behaviour of the host is part of the extended phenotype of a parasite (Andersen et al., 2009), thus having evolutionary significance for the latter. In the so-called “zombie ants”, the fungus induces a set of actions that differs from the normal habits of those insects (including erratic wandering, convulsions preventing the host to climb high on trees, and death grips on specific points on leaves), which are function to bring the host to a suitable location that maximises the spread of fungal spores (Hughes et al., 2011).
This kind of interaction is also present in other fungi-arthropods symbiotic relationships, sometimes even attesting for convergent evolution in very distantly related fungi parasitizing different hosts (Hughes et al., 2016). Following Tinbergen’s (1963) view on understanding animal behaviour, there are four different approaches to the study of the manipulations carried out by the parasites. Those approaches focus on: 1) the functionality of the modified behaviour, 2) its phylogenetic history, 3) its proximate causes, and 4) its ontogenetic development.
We can discuss the CBI infected under the point of view of at least the first and the last approaches, intertwining behavioural aspects with morphological changes caused by Cordyceps. It is known that researchers in the TLoU universe were investigating the proximate causes behind the Cordyceps-human interactions (i.e., the molecular and physical mechanisms that acted on the host, generating the manifestations of the infection) to better understand the infection and develop either a cure or a vaccine. This is not our focus here (also because we, fortunately, do not have specimens of infected available), but we can briefly report that the CBI fungus operates starting from the victim’s brain, manipulating its nervous system and its perceptions. More precise information on the molecular path behind this are not available to us. We cannot say much about the phylogenetic approach either, due to the unknown specific identity of the CBI fungus and the obvious impossibility of obtaining molecular sequences useful to reconstruct its phylogeny.
From another point of view, the progression of the CBI has a clear ontogenetic aspect, related to the development of the parasitoid fungus inside (and outside) its host. This is manifested in the very different appearances of the infected at subsequent stages and also in the behaviours displayed. Early stages of the infection seem to be more mobile, forming large hordes that move for long distances in a sort of dispersal event (maybe even a migration if the movement from and towards the areas involved is persistent; Dingle & Drake, 2007). Participation of stalkers in hordes, even though not clearly observed by us, is expected given their mobility; possibly, their presence in such gatherings is simply overlooked because of their more secretive habits. Later CBI stages, such as the shamblers and the bloaters, seem to lose their mobility, becoming more restricted to limited areas that are often those where fructification of other infected takes place.
Development of fungal structures over the body of the infected is another ontogenetical manifestation of the CBI, being logically more expressed in later stages. In this context, a peculiar feature is the development of echolocation abilities and the related breakage of the host’s skull. Echolocation starts to appear in stage 2 infected (stalkers), but it is still imperfect. It develops further in clickers, when the fungus opens a way through the skull of the host, likely creating a structure that improves the echolocation ability. Bloaters are good echolocators as well. It is impossible to say whether the structure emerging in clickers evolved to favour echolocation or if the development of echolocation is a by-product of this morphological trait originated to counterbalance the loss of other senses. It is interesting to note that shamblers do not present the same degree of breakage in their skull as clickers and bloaters do, in spite of being considered a possible alternative stage 4 of the infection. If this hypothesis were true, then one would expect the development of the fungus through the cranial bones to remain quite evident, given that the shambler would derive from a clicker displaying this damage. A possible explanation for the different morphology seen in these infected could be that they indeed represent a parallel line of development for the Cordyceps, but coming from an earlier stage (i.e., either runners or stalkers) and accelerating the realization of a bloater-like morphology for some reason. Unfortunately, there is no way to verify this hypothesis based on our current knowledge of TLoU infected, and we can only speculate for the moment. In all those ontogenetic aspects of the infection, the Rat King is of course an extreme due to its peculiar nature and the uniquely extended time of fungal development.
Adding to the previously-mentioned traits, another important ontogeny-related feature in the CBI is the timing of fructification. Considering what we can see in TLoU and TLoU2, it seems that fructification takes place when the infected is either at stage 2 (stalkers) or stage 3 (clickers) of the infection. Given that passage from the stalker to the clicker phase happens roughly around one year from contamination, this means that it is apparently possible for the Cordyceps to reach maturity in less than one year, and that either maturation or fructification time may be variable or delayable. Factors related to this postponed maturation/fructification are unknown, but they may include availability of a place suitable for spore release. We are not aware of any fructification event in either shamblers or bloaters, even though this is not definitive evidence of a lack of fructification abilities in Cordyceps at those stages of the infection (absence of evidence is not evidence of absence, in particular when we have so little data to work with).
All the morphological and behavioural changes Cordyceps induces in the hosts can be observed under a functional lens as well. Within its life cycle, the main “objective” of the Cordyceps fungus that causes the CBI is to reproduce, that is, to produce spores and spread the infection to other hosts. There are numerous factors that play a role in this, including: 1) availability of possible new hosts (i.e., uninfected humans); 2) capacity of the infected to actually infect potential new hosts; 3) possibility of the infected and spores to survive long enough to reach maturation; and 4) presence of suitable environments for fructification and spore survival. All modifications induced in the host by the fungus likely take part in trying to secure success for its main “objective”.
The progressive enhancement in strength and damage resistance that is observed throughout different infection stages clearly gives an advantage in the struggles with possible new hosts, but also with other types of prey. It is demonstrated that the infected attack other animals as well, which are likely not susceptible to the CBI and so, that is done maybe for food and sustenance. Alternatively, these attacks may be simple by-products of the increased aggression of the infected, even though observations of infected actively consuming the meat of the hunted preys may suggest otherwise. In any case, the strengthening of the host implemented by the fungus works in parallel to a decrease in its response to pain and in tandem with increased ferocity and lack of self-preservation and fleeing; all of which enhances the chances of CBI spreading. Suppression of host responses to external stimuli has already been observed in other parasitoid fungi (de Bekker et al., 2015), being thus not surprising in this case. Parasitoid fungi infecting ants are also known to remodel the cuticle of the insect to increase resistance of their weaker parts (Anderson et al., 2009).
The search for new hosts may be invoked to explain the formation of hordes that travel across great distances in early CBI stages. Infected that form hordes are the ones in which CBI transmission is documented through active aggression towards a potential new host and also those in which development of the fungus to fructification is still undisputedly possible. Later, less mobile stages are not documented to bite when they attack, thus not spreading the CBI by fluid contact. As mentioned above, they are also not known to fructificate, even though this may be biased by lack of observations.
These behavioural differences may be due to different roles in the Cordyceps life cycle of the two groups of infected. Early stages, up to the clickers, may be devoted to spreading the infection by chasing and attacking suitable hosts as well as moving towards new areas. On the other hand, later stages may have a protective role: being tied to the places where fructification of other infected takes place, they may act as defenders, confronting and killing whoever threatens the defenceless fully-developed Cordyceps.
Indeed, humans actively destroy fructification clusters in TLoU to clean the areas where they are located. Adaptations devoted to host protection seems to be present in real Ophiocordyceps, but are generally focused on protecting the infected ants by moving it away from sources of harm or producing antimicrobial compounds (Anderson et al., 2009), but not protecting other parasitized ants. Under this point of view, the behaviour displayed by CBI infected looks more like nest protection, similar to social insects. Of course, we cannot confirm the real nature of the behaviours observed in shamblers and bloaters, and they may alternatively result simply from an extreme development of the prolonged infection causing very high aggressiveness but making it impossible for them to bite. This, however, would not explain why these infected progressed so much without fructification; so, a different (and possibly functional) reason may be involved. The protection role hypothesis is further supported by the development of offensive weapons: i.e., the ichor-filled rounded structures.
CONCLUSIONS
Looking at characters from videogames and other fictional contexts with a natural science approach is not easy and inherently subject to a high degree of speculative thinking, due to the general limitation of the observations that can be made together with the impossibility of actively confirming hypotheses derived from the data collected. Still, highlighting and discussing such data remains an interesting exercise for those who are interested in diving deeper into the lore of works of fiction that fascinated them.
Despite being of course not canonical, the hypotheses proposed here to explain the observed phenomena contribute to pointing out emergent narrative elements that may or may not be in the original concept created by the authors. In this context, CBI infected from the TLoU game series are an excellent example. Features that can be observed in the different kinds of infected were of course included by the developers because they are functionally tuned to the need of an action-adventure game or to their integration within the setting of the series. As an example, progressive stages displaying increasing strength and resistance, as well as the development of new abilities, clearly respond to the requirement of presenting the players with new challenges while they advance in the game, whereas fungal structures appearing on the infected bodies help in depicting them as victims of the Cordyceps parasitism. However, all these observed features can still be coherently interpreted using a “scientific” eye, being thus both the umpteenth evidence of the impressive level of attention to details in Naughty Dog’s work and an opportunity for people like us to further explore the universe they created.
As such, a careful description of the morphology and behaviour of the infected brought us to the reconstruction of an ontogenetical pattern in most of the observed features, which parallels the idea of an increased difficulty in dealing with them in some cases, while in other cases it revealed interesting departures from a simple continuous development. The most outstanding examples of the latter are probably the apparent ability of some infected to avoid fructification and progress to a successive infection stage and maybe even more the possibly unclear position of shamblers within the standard CBI progression. Both in the case of shamblers being an alternative form to bloaters (appearing because of more humid conditions) and the case of them being an earlier-diverging form, their existence opens up tantalizing scenarios about the adaptive potential of the Cordyceps causing the CBI and the evolutionary outcomes that it can have. Similar implications may be retrieved from the functional analysis of the reported data, with particular emphasis on the recognition of two possible groups within the CBI stages. These groups may play very different roles in the Cordyceps life cycle: one group is composed by runners, stalkers, and clickers, having the “task” of spreading the infection; the other is composed by shamblers and bloaters, having a “nest”-protection role. Presence of anomalies such as the Rat King further stresses the adaptability of the CBI Cordyceps as well as the complex nature of the interactions within this, its hosts, and the surrounding environment. Further insights in all these issues may be fuelled by the prospective publication of further TLoU material in the future, such as (but hopefully not only) the forthcoming TV series.
REFERENCES
Andersen, S.B.; Gerritsma, S.; Yusah, K.M.; Mayntz, D.; Hywel-Jones, N.L.; Billen, J.; Boomsma, J.J.; Hughes, D.P. (2009) The life of a dead ant: the expression of an adaptive extended phenotype. The American Naturalist 174(3): 424–433.
de Bekker, C.; Ohm, RA.; Loreto, R.G.; Sebastian, A.; Albert, I.; Merrow, M.; Brachmann, A.; Hughes, D.P. (2015) Gene expression during zombie ant biting behavior reflects the complexity underlying fungal parasitic behavioral manipulation. BMC Genomics 16: 620.
Dingle, H. & Drake, V.A. (2007) What is migration? BioScience 57(2): 113–121.
Hughes, D.P; Andersen, S.B.; Hywel-Jones, N.L.; Himaman, W.; Billen, J.; Boomsma, J.J. (2011) Behavioral mechanisms and morphological symptoms of zombie ants dying from fungal infection. BMC Ecology 11: 13.
Hughes, D.P.; Araújo, J.; Loreto, R.; Quevillon, L.; de Bekker, C.; Evans, H.C. (2016) From so simple a beginning: the evolution of behavioral manipulation by fungi. Advances in Genetics 94: 437–469.
Tinbergen, N. (1963) On aims and methods of ethology. Zeitschrift für Tierpsychologie 20(4): 410–433.
Turi, T. (2019) Neil Druckmann discusses new The Last of Us Part II details. PlayStation Blog. Available from: https://blog.playstation.com/2019/10/09/neil-druckmann-discusses-new-the-last-of-us-part-ii-details/?ref-tag=252542 (Date of access: 02/Dec/2021).
ACKNOWLEDGMENTS
We would like to give our heartfelt thanks to all the people at Naughty Dog, for developing both TLoU games and by doing so, allowing us to live the amazing experience that was playing them. Our gratitude also goes to the whole community of the TLoU Fandom Wiki, whose fine work was a great help for refining and filling gaps in our personal observations. Having such a detailed and precise resource at hand was of utmost importance. We further thank Flavia Strani for the amazing artwork she made for our Fig. 1, and the editor-in-chief of the Journal of Geek Studies, Rodrigo B. Salvador, for his assistance while preparing this manuscript.
About the authors
Andrea Villa is a vertebrate palaeontologist, studying the comparative osteology and palaeodiversity of amphibians and reptiles. He is deeply passionate about nature, videogames, science fiction and, you know, all that kind of stuff…
Alessio del Negro carries out business consultancy activities in order to assess the processes of a company as well as efficacy of its internal control. He has a great passion for videogames, TV series and boardgames, and often gets drawn into “stranger things” by an even stranger vertebrate palaeontologist.