Skip to content
RSS Feed
Bluesky
Type your email…
Subscribe
Search
Journal of Geek Studies
About
Guidelines for authors
Current Issue
Archives
Vol. 13(1): Jun/2026
Vol. 12(2): Dec/2025
Vol. 12(1): Jun/2025
Vol. 11(2): Dec/2024
Vol. 11(1): Jun/2024
Older issues
Editorial Team
Partners
Contact
Zoology
July 19, 2026
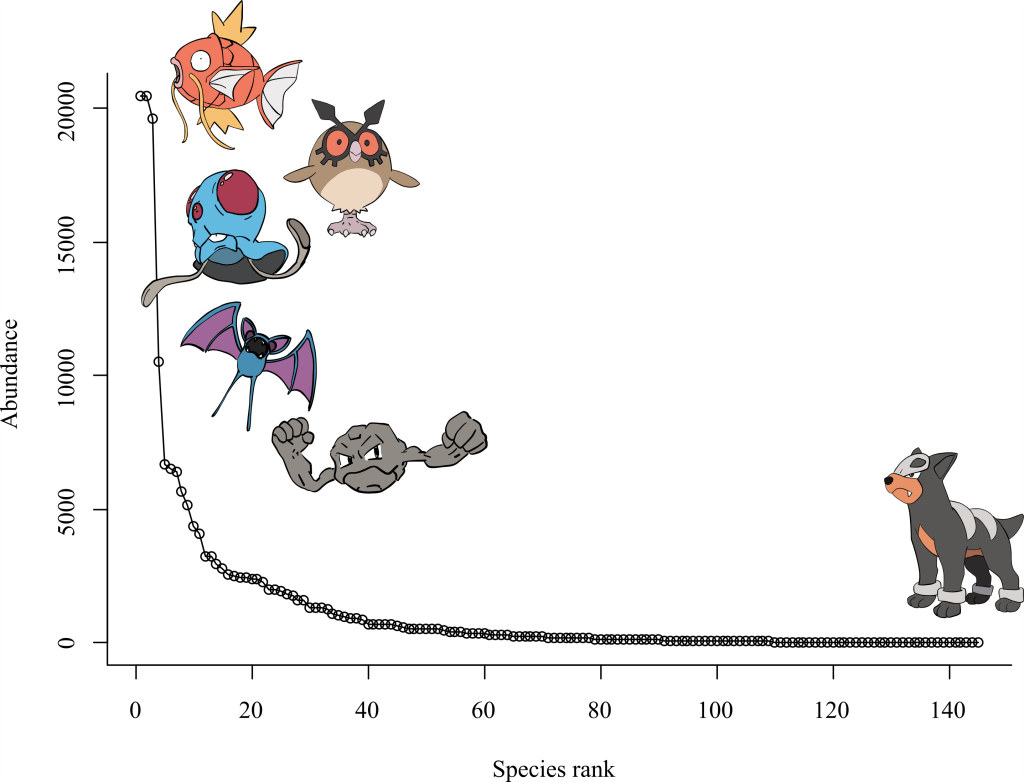
Ecological patterns in Pokémon games reflect those observed in the natural world
Biology
,
Video Games
Ecology
,
Pokémon
,
Zoology
May 1, 2026
A kawaii slug? The sea bunny Jorunna parva in pop culture
Anime & Manga
,
Biology
,
Cinema
,
Japan Studies
Ecology
,
Malacology
,
Marine Biology
,
Nature Conservation
,
Taxonomy
,
Zoology
February 5, 2026
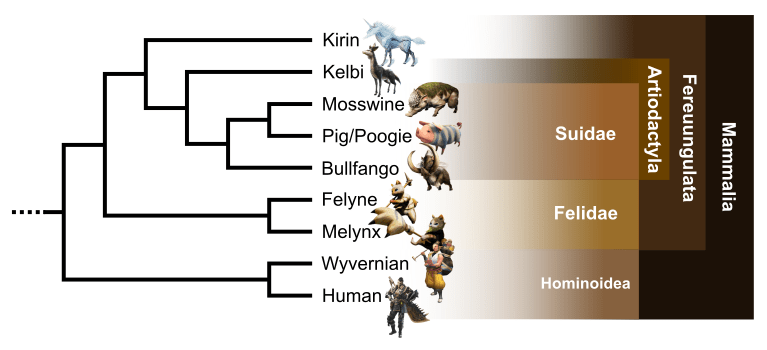
Monster Hunter: an evolutionary story
Biology
,
Video Games
,
Zoology
Monster Hunter
,
Taxonomy
,
Zoology
November 11, 2025
Can conservation really be effective through video games? A case study on Jack Barau and some perspectives
Biology
,
Education
,
Environmental Sciences
,
Video Games
,
Zoology
Ecology
,
Game Development
,
Nature Conservation
,
Ornithology
,
Zoology
November 1, 2025
When trash stops being a treasure: pollution and its effects in Another Crab’s Treasure
Biology
,
Environmental Sciences
,
Video Games
Carcinology
,
Ecology
,
Marine Biology
,
Nature Conservation
,
Zoology
August 24, 2025
Molluscs of the Multiverse: molluscan diversity in Magic: The Gathering
Biology
,
Mollusks
,
Tabletop Games
Magic the Gathering
,
Malacology
,
Taxonomy
,
Trading Card Games
,
Zoology
August 24, 2024
Euarthropod diversity in Pokémon: searching for the ancestral type
Biology
,
Video Games
,
Zoology
Arthropodology
,
Pokémon
,
Taxonomy
,
Zoology
July 8, 2024
What’s the deal with the blue “robins” in gacha games?
Biology
,
Video Games
Birdwatching
,
Fate Grand Order
,
Gacha
,
Honkai Star Rail
,
Mobile Games
,
Ornithology
,
Zoology
June 8, 2024
Rewriting Earth: environmentally-minded comics to move all audiences
Biology
,
Comics
,
Interview
Ecology
,
Education
,
Nature Conservation
,
Webcomics
,
Zoology
November 11, 2023
My neighbor Linnaeus: The influence of Studio Ghibli in zoological nomenclature
Anime & Manga
,
Biology
,
Cinema
,
Japan Studies
,
Zoology
Ecology
,
Fantastic Beasts
,
Ghibli
,
Taxonomy
,
Zoology
1
2
3
…
5
Next Page
Subscribe
Subscribed
Journal of Geek Studies
Join 309 other subscribers
Sign me up
Already have a WordPress.com account?
Log in now.
Journal of Geek Studies
Subscribe
Subscribed
Sign up
Log in
Report this content
View site in Reader
Manage subscriptions
Collapse this bar