Skip to content
RSS Feed
Bluesky
Type your email…
Subscribe
Search
Journal of Geek Studies
About
Guidelines for authors
Current Issue
Archives
Vol. 13(1): Jun/2026
Vol. 12(2): Dec/2025
Vol. 12(1): Jun/2025
Vol. 11(2): Dec/2024
Vol. 11(1): Jun/2024
Older issues
Editorial Team
Partners
Contact
Taxonomy
May 1, 2026
A kawaii slug? The sea bunny Jorunna parva in pop culture
Anime & Manga
,
Biology
,
Cinema
,
Japan Studies
Ecology
,
Malacology
,
Marine Biology
,
Nature Conservation
,
Taxonomy
,
Zoology
February 5, 2026
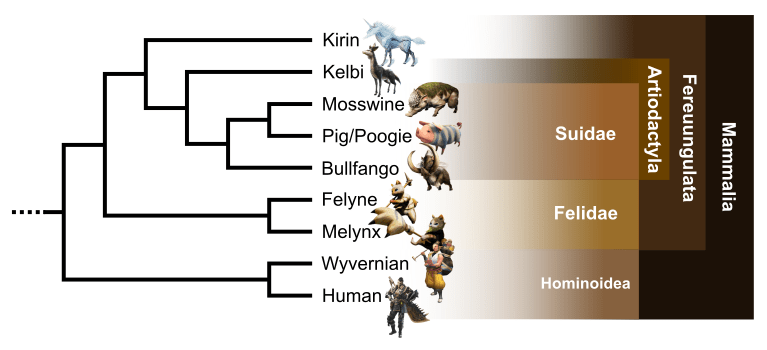
Monster Hunter: an evolutionary story
Biology
,
Video Games
,
Zoology
Monster Hunter
,
Taxonomy
,
Zoology
August 24, 2025
Molluscs of the Multiverse: molluscan diversity in Magic: The Gathering
Biology
,
Mollusks
,
Tabletop Games
Magic the Gathering
,
Malacology
,
Taxonomy
,
Trading Card Games
,
Zoology
December 2, 2024
Xenomorphia ex machina: the zoology and biochemistry of xenomorphs from the Alien franchise
Biology
,
Cinema
,
Horror
,
Zoology
Alien
,
Fantastic Beasts
,
Horror Movies / Books
,
Science Fiction
,
Taxonomy
,
Xenomorphs
August 24, 2024
Euarthropod diversity in Pokémon: searching for the ancestral type
Biology
,
Video Games
,
Zoology
Arthropodology
,
Pokémon
,
Taxonomy
,
Zoology
November 11, 2023
My neighbor Linnaeus: The influence of Studio Ghibli in zoological nomenclature
Anime & Manga
,
Biology
,
Cinema
,
Japan Studies
,
Zoology
Ecology
,
Fantastic Beasts
,
Ghibli
,
Taxonomy
,
Zoology
September 2, 2023
Dark wings, bright insights: a comprehensive analysis of corvid species in Pokémon games
Biology
,
Video Games
,
Zoology
Birdwatching
,
Ornithology
,
Pokémon
,
Taxonomy
,
Zoology
April 15, 2023
The lure of the deep sea: anglerfish as movie monsters
Animation
,
Biology
,
Cinema
Ichthyology
,
Marine Biology
,
Star Wars
,
Taxonomy
October 13, 2022
Cicadas in Japanese video games and anime
Anime & Manga
,
Biology
,
Japan Studies
,
Literature
Ecology
,
Entomology
,
Fate Grand Order
,
Higurashi
,
Pokémon
,
Taxonomy
,
Zoology
February 26, 2022
A critical reappraisal of dinosaur reconstructions in Zoo Tycoon 2: Extinct Animals
Archaeology
,
Biology
,
Paleontology
,
Video Games
Ecology
,
Nature Conservation
,
Simulation video game
,
Taxonomy
,
Zoology
1
2
3
Next Page
Subscribe
Subscribed
Journal of Geek Studies
Join 309 other subscribers
Sign me up
Already have a WordPress.com account?
Log in now.
Journal of Geek Studies
Subscribe
Subscribed
Sign up
Log in
Report this content
View site in Reader
Manage subscriptions
Collapse this bar