Skip to content
RSS Feed
Bluesky
Type your email…
Subscribe
Search
Journal of Geek Studies
About
Guidelines for authors
Current Issue
Archives
Vol. 13(1): Jun/2026
Vol. 12(2): Dec/2025
Vol. 12(1): Jun/2025
Vol. 11(2): Dec/2024
Vol. 11(1): Jun/2024
Older issues
Editorial Team
Partners
Contact
March 8, 2025
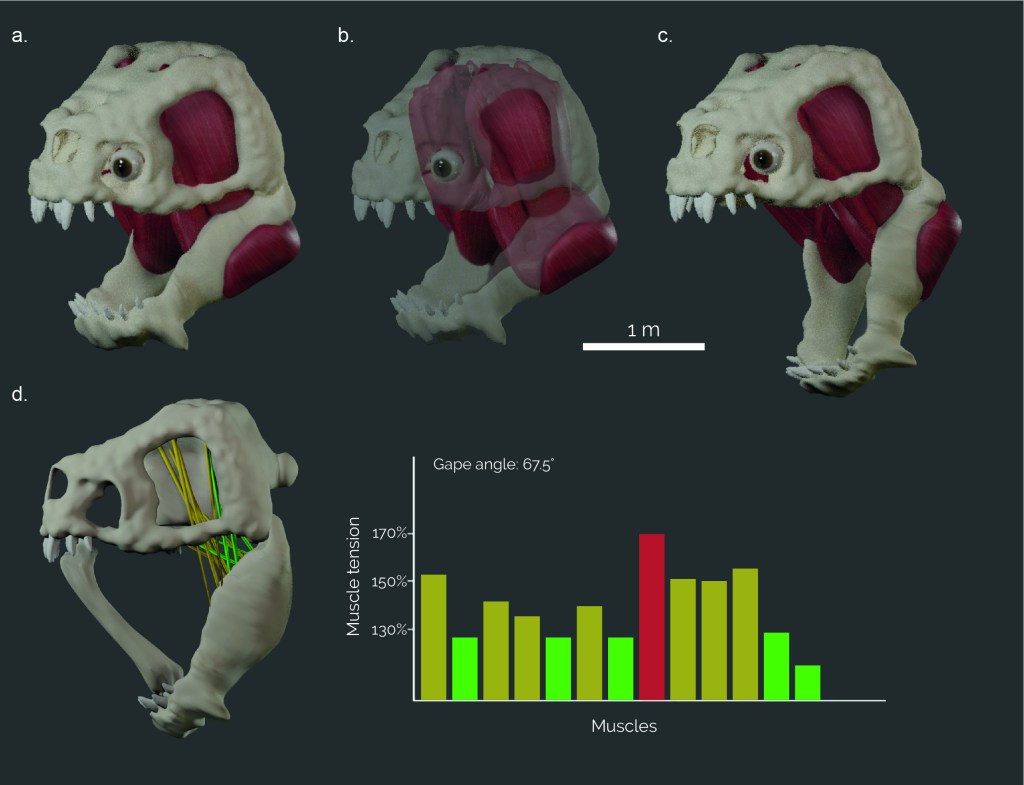
Is a bone a viable weapon when combating a Rancor? Estimating the bite force of an intergalactic mega-predator
Biology
,
Cinema
,
Paleontology
,
Physics
,
Television Series
Dynamics
,
Fantastic Beasts
,
Finite element method
,
Science Fiction
,
Star Wars
February 26, 2022
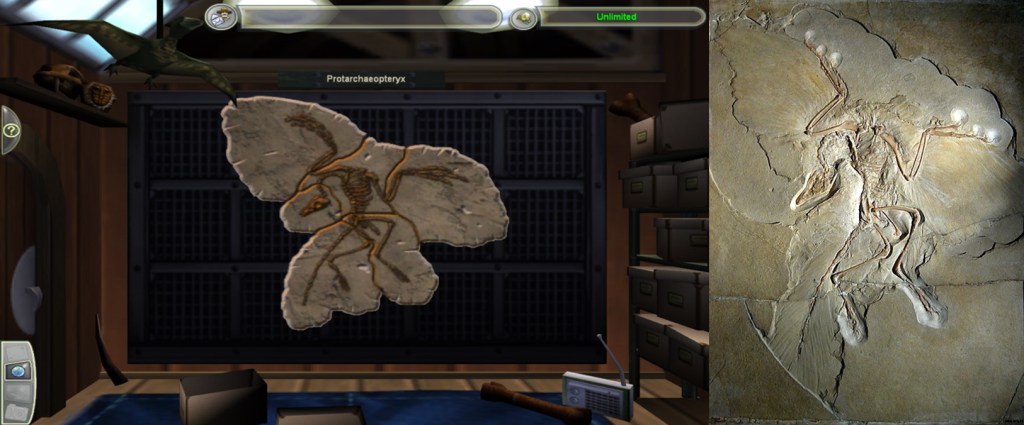
A critical reappraisal of dinosaur reconstructions in Zoo Tycoon 2: Extinct Animals
Archaeology
,
Biology
,
Paleontology
,
Video Games
Ecology
,
Nature Conservation
,
Simulation video game
,
Taxonomy
,
Zoology
August 5, 2021
Short note on recent sightings of Rodan in Godzilla Singular Point
Anime & Manga
,
Biology
,
Cinema
,
Paleontology
Ecology
,
Fantastic Beasts
,
Giant Monsters
,
Godzilla
,
Taxonomy
,
Zoology
July 5, 2020
The One Born of Fire: a pterosaurological analysis of Rodan
Biology
,
Cinema
,
Paleontology
Ecology
,
Giant Monsters
,
Godzilla
,
Taxonomy
,
Trending
,
Zoology
March 22, 2020
Fossil Pokémon and the foibles of Paleontology
Biology
,
Paleontology
,
Video Games
Paleontology
,
Pokémon
February 11, 2020
A paleontological outlook on the Super Mario Bros. movie
Biology
,
Cinema
,
Paleontology
,
Video Games
Dinosaurs
,
Super Mario
October 30, 2019
Mondo Museum: a sim game to build your own world-class dream museum
Archaeology
,
Biology
,
History
,
Paleontology
,
Video Games
Indie Games
,
Museum
,
Simulation video game
November 4, 2018
Moa v Superman
Biology
,
Comics
,
Paleontology
,
Super Heroes
DC Comics
,
Ornithology
,
Superman
,
Taxonomy
,
Zoology
June 3, 2018
One squid to rule them all
Biology
,
Literature
,
Mollusks
,
Paleontology
,
Video Games
Lord of the Rings
,
Taxonomy
,
Zoology
Subscribe
Subscribed
Journal of Geek Studies
Join 309 other subscribers
Sign me up
Already have a WordPress.com account?
Log in now.
Journal of Geek Studies
Subscribe
Subscribed
Sign up
Log in
Report this content
View site in Reader
Manage subscriptions
Collapse this bar