Skip to content
RSS Feed
Bluesky
Type your email…
Subscribe
Search
Journal of Geek Studies
About
Guidelines for authors
Current Issue
Archives
Vol. 13(1): Jun/2026
Vol. 12(2): Dec/2025
Vol. 12(1): Jun/2025
Vol. 11(2): Dec/2024
Vol. 11(1): Jun/2024
Older issues
Editorial Team
Partners
Contact
Pokémon
July 19, 2026
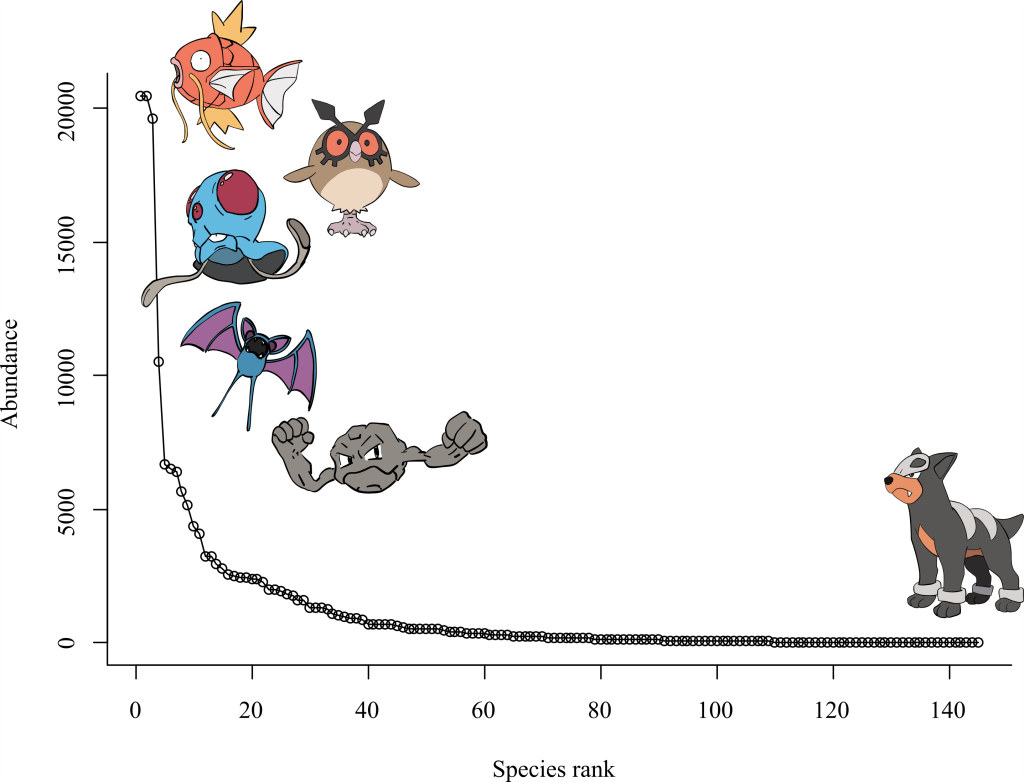
Ecological patterns in Pokémon games reflect those observed in the natural world
Biology
,
Video Games
Ecology
,
Pokémon
,
Zoology
July 11, 2025
Predicting competitive Pokémon VGC leads using Latent Semantic Analysis: a data-driven approach to team matchups
Artificial Inteligence
,
Simulation
,
Video Games
Natural language processing
,
Pokémon
May 1, 2025
Pokérus: unraveling the biology of the Pokémon virus
Biology
,
Video Games
Pokémon
,
Virology
August 24, 2024
Euarthropod diversity in Pokémon: searching for the ancestral type
Biology
,
Video Games
,
Zoology
Arthropodology
,
Pokémon
,
Taxonomy
,
Zoology
September 2, 2023
Dark wings, bright insights: a comprehensive analysis of corvid species in Pokémon games
Biology
,
Video Games
,
Zoology
Birdwatching
,
Ornithology
,
Pokémon
,
Taxonomy
,
Zoology
June 26, 2023
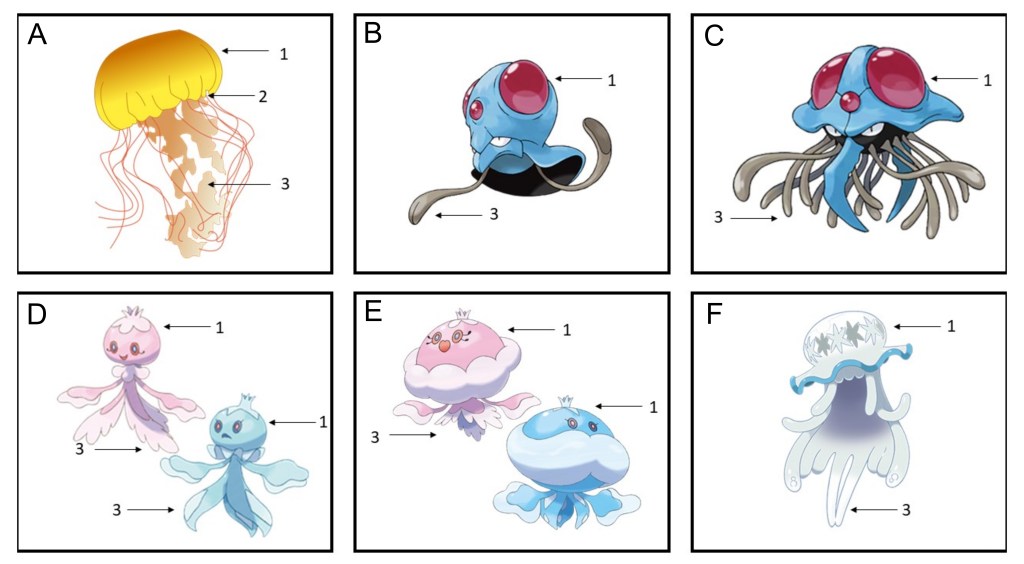
From reality to fiction: cnidarians that inspire the Pokémon world
Biology
,
Video Games
,
Zoology
Cnidaria
,
Ecology
,
Marine Biology
,
Morphology
,
Pokémon
,
Zoology
October 13, 2022
Cicadas in Japanese video games and anime
Anime & Manga
,
Biology
,
Japan Studies
,
Literature
Ecology
,
Entomology
,
Fate Grand Order
,
Higurashi
,
Pokémon
,
Taxonomy
,
Zoology
July 18, 2021
Floral gigantism in the Pokémon world
Anime & Manga
,
Biology
,
Uncategorized
,
Video Games
Botany
,
Pokémon
August 20, 2020
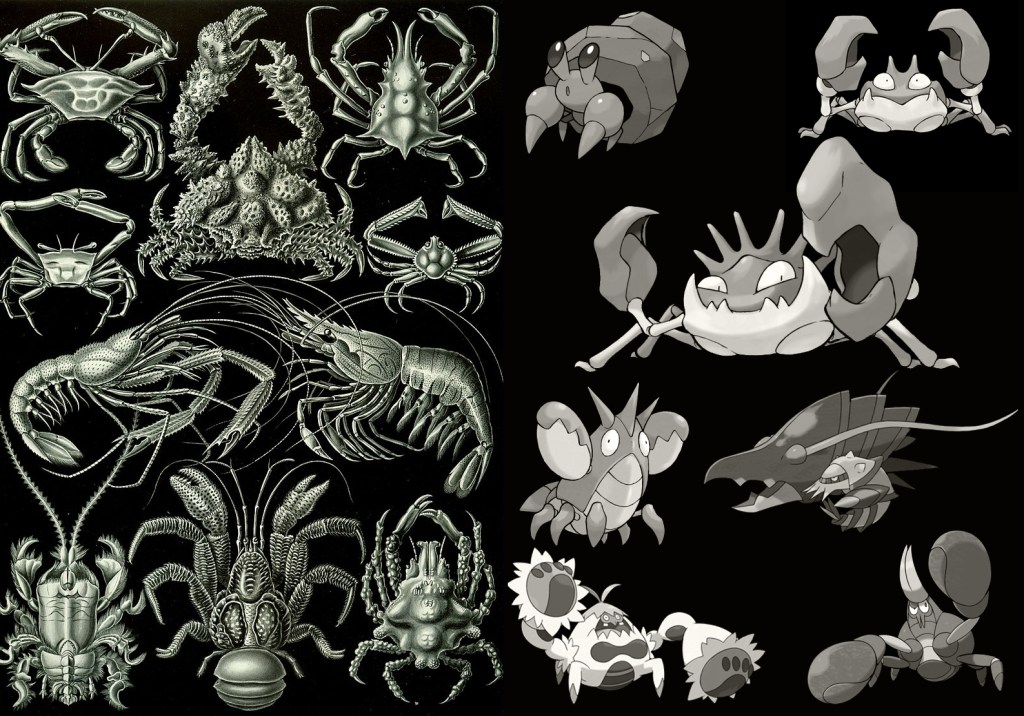
Pokécrustacea: the crustacean-inspired Pokémon
Biology
,
Video Games
,
Zoology
Carcinology
,
Marine Biology
,
Pokémon
,
Zoology
June 24, 2020
What’s your favourite Pokémon? Pocket monster popularity reflects interest in real-world Biology
Biology
,
Video Games
,
Zoology
Ecology
,
Pokémon
,
Survey
,
Zoology
1
2
3
Next Page
Subscribe
Subscribed
Journal of Geek Studies
Join 309 other subscribers
Sign me up
Already have a WordPress.com account?
Log in now.
Journal of Geek Studies
Subscribe
Subscribed
Sign up
Log in
Report this content
View site in Reader
Manage subscriptions
Collapse this bar