Augusto B. Mendes1, Felipe V. Guimarães2, Clara B. P. Eirado-Silva1 & Edson P. Silva1
1Universidade Federal Fluminense, Niterói, RJ, Brazil.
2Universidade do Estado do Rio de Janeiro, São Gonçalo, RJ, Brazil.
Emails: augustobarrosmendes (at) yahoo (dot) com (dot) br; felipevieiragui (at) gmail (dot) com; clara.eirado (at) gmail (dot) com; gbmedson (at) vm (dot) uff (dot) br
https://doi.org/10.5281/zenodo.10644563
Pokémon, or Pocket Monsters, was originally created for videogames, becoming a worldwide fever among kids and teenagers in the end of the 1990’s and early 2000’s. Currently, it is still a success, with numerous games, a TV series, comic books, movies, a Trading Card Game, toys and collectibles. Through its core products and vibrant merchandising, Pokémon took over the world, influencing pop culture wherever it landed. Despite losing some steam in the early 2010’s, Pokémon is now back to its previous uproar with the release of Pokémon GO, an augmented reality (AR) game for smartphones. This game launched in 2016, with almost 21 million users downloading it in the very first week in the United States alone (Dorward et al., 2017). Thus, Pokémon is indubitably an icon in pop culture (Schlesinger, 1999a; Tobin, 2004).
The origin of Pokémon goes back to two role-playing video games (created by Satoshi Tajiri and released by Nintendo for the Game Boy; Kent, 2001): Pokémon Green and Pokémon Red, released in Japan in 1996. In the West, the Green version never saw the light of day, but the Red and Blue versions were released in 1998, selling together more than 10 million copies. Also in 1998, the Yellow version of the game was released, which has as its most distinct feature the possibility of having Pikachu (the most famous Pokémon) walking side by side with the player in the game. Pokémon Green, Red, Blue and Yellow are the so-called “first generation” of games in the franchise. Today, the Pokémon series is in its seventh generation, with 29 main games released, besides several spin-offs. The TV series, on the other hand, is in its sixth season, with more than 900 episodes.
The games and TV series take place in regions inhabited by many Pokémon and humans. The mission of the protagonist is to win competitions (“Pokémon battles”) against gym leaders who are spread across different cities and regions. For each victory, the protagonist receives a gym badge; with eight badges, he/she is allowed to enter the Pokémon League to try and become the Champion.
For each generation, new Pokémon (and an entire new region) are introduced. In this way, the creatures have a homeland, although most can appear in other regions as well (Schlesinger, 1999b; Whitehill et al., 2016). The seven main regions are: Kanto, Johto, Hoenn, Sinnoh, Unova, Kalos and Alola.
In every region, there are numbered routes that connect cities and landmarks and in which the protagonist travels, finding the monsters in their natural habitats and interacting with other characters. These routes comprise a great range of environments, such as forests, caves, deserts, mountains, fields, seas, beaches, underwater places, mangroves, rivers and marshes, which usually display a huge diversity of Pokémon.
In addition to winning the Pokémon League, the protagonist must complete the Pokédex, a digital encyclopedia of Pokémon. In other words, the trainer must catch all the Pokémon that live in that region, registering each capture in the Pokédex. Each Pokémon has a registry number and an entry text in the Pokédex. Pokémon are usually found in nature, and may be captured with a device called “Pokéball”. Pokéballs are small enough to fit in a pocket, hence the name “Pocket Monsters” (Whitehill et al., 2016).
NOT AS MONSTRUOUS AS WE THINK
In the world depicted in the games, there are 801 Pokémon, belonging to one or two of the following 18 types: Normal, Fire, Fighting, Water, Flying, Grass, Poison, Electric, Ground, Psychic, Rock, Ice, Bug, Dragon, Ghost, Dark, Steel and Fairy (Bulbapedia, 2017). Almost all Pokémon are based on animal species, some of them are based on plants or mythological creatures, and a few are based on objects. Curiously, all Pokémon are oviparous, which means they all lay eggs (their development happens inside of an egg and outside of their mother’s body); of course, in the real natural world, this is a reproductive strategy of animals such as fishes, amphibians, reptiles, birds and many kinds of invertebrates (Blackburn, 1999). Moreover, Pokémon might “evolve”, usually meaning they undergo some cosmetic changes, become larger and gain new powers.
In the present work, the Pokémon world was approached by analogies with the real natural world, establishing parallels with actual animals.
A remarkable group of animals represented in Pokémon is the fishes. Fishes are the largest group of vertebrates, with more than 32,000 species inhabiting marine and freshwater environments, a number that roughly corresponds to half of all described vertebrates (Nelson et al., 2016). Showing ample morphological and behavioral variety and living in most of the aquatic ecosystems of the planet, fishes are well represented in the Pokémon world, therefore offering a great opportunity for establishing parallels between the two worlds. The creators of the games not only used the morphology of real animals as a source of inspiration for the monsters, but also their ecology and behavior.
Based on these obvious connections between real fishes and Pokémon, the aim of this work is to describe the ichthyological diversity found in Pokémon based on taxonomic criteria of the classification of real fishes. Ultimately, our goal is to offer useful material for both teaching and the popularization of science.
Table 1. Taxonomic classification of the fish Pokémon. Abbreviations: Ch = Chondrichthyes; Gn = Gnathostomata; Pe = Petromyzontomorphi; Pt = Petromyzontida; Os = Osteichthyes. All images obtained from The Official Pokémon Website (2016).

GOTTA CATCH ‘EM FISHES!
The first step of our research was a search in the Pokédex (The Official Pokémon Website, 2016) for Pokémon which were related to fishes. The criterion used was the Pokémon’s morphology (resemblance to real fishes). Afterwards, the “fish Pokémon” were classified to the lowest taxonomic level (preferably species, but when not possible, genus, family or even order).
This classification of the Pokémon allowed the comparison of biological data (such as ecological, ethological, morphological traits) from Bulbapedia (2017) with the current knowledge on real fishes (e.g., Nelson et al., 2016). Bulbapedia is a digital community-driven encyclopedia created in 2004 and is the most complete source regarding the pocket monsters.
The final step was a search in online scientific databases (Fishbase, Froese & Pauly, 2016; and Catalog of Fishes, Eschmeyer et al., 2016) in order to obtain the current and precise taxonomy and additional information on habitats, ecology etc. of the fish species.
In the present work, the taxonomic classification used was that proposed by Nelson et al. (2016), who consider the superclasses Petromyzontomorphi (which includes the class Petromyzontida, that is, the lampreys) and Gnathostomata (the jawed vertebrates). Gnathostomata, in turn, includes the classes Chondrichthyes (cartilaginous fishes) and Osteichthyes (bony fishes). Along with this classification, we used the classification proposed by the database ITIS (Integrated Taxonomic Information System, 2016) for comparison at all taxonomic levels. Following identification, the “fish Pokémon” were described regarding their taxonomic and ecological diversity.
POCKET FISHES
As a result of our search, 34 fish Pokémon were identified (circa 4% of the total 801 Pokémon; Table 1) and allocated in two superclasses, three classes, eighteen orders, twenty families and twenty-two genera. Eighteen of the 34 fish Pokémon (circa 53%) could be identified to the species level (Table 2). The features of the real fishes which probably inspired the creation of the Pokémon and other relevant information are described below for each species. To enrich the comparisons, images of the Pokémon (obtained from the Pokédex of The Official Pokémon Website; http://www.pokemon.com) and of the real fishes (illustrations by one of us, C.B.P. Eirado-Silva) follow the descriptions.
Table 2. Taxonomic diversity of the fish Pokémon.

Horsea and Seadra
Species: Hippocampus sp.; Common name: seahorse.
The Pokémon Horsea and Seadra (Fig. 1), which debuted in the first generation of the franchise, were based on seahorses. The long snout, ending in a toothless mouth (Foster & Vincent, 2004), the prehensile, curved tail (Rosa et al., 2006) and the salient abdomen are features of the real fishes present in these Pokémon. Seahorses belong to the genus Hippocampus, presently composed of 54 species (Nelson et al., 2016). The males have a pouch in their bellies where up to 1,000 eggs are deposited by the females. In this pouch, the eggs are fertilized and incubated for a period ranging from 9 to 45 days (Foster & Vincent, 2004). Due to overfishing for medicinal and ornamental purposes, as well habitat destruction, about 33 species of seahorses are considered threatened (Rosa et al., 2007, Castro et al., 2008; Kasapoglu & Duzgunes, 2014).

Figure 1. Horsea, Seadra and Hippocampus sp.
Goldeen and Seaking
Species: Carassius auratus; Common name: goldfish.
Goldeen and Seaking (Fig. 2) were based on the goldfish. This species is one of the most common ornamental fishes worldwide (Soares et al., 2000; Moreira et al., 2011) and it is widely used in studies of physiology and reproduction due to its docile behavior and easy acclimatization to artificial conditions (Bittencourt et al., 2012; Braga et al., 2016). The resemblance between the goldfish and the Pokémon include morphological features, such as the orange/reddish color and the long merged fins, and the name “Goldeen”. The name Seaking, on the other hand, may be a reference to another common name of the species, “kinguio”, from the Japanese “kin-yu” (Ortega-Salas & Reyes-Bustamante, 2006).

Figure 2. Goldeen, Seaking and Carassius auratus.
Magikarp
Species: Cyprinus carpio; Common name: common carp.
Possibly the most famous fish Pokémon, Magikarp (Fig. 3) was based on a common carp, a species present in Europe, Africa and Asia, widely used in pisciculture due to its extremely easy acclimatization to many freshwater environments and the high nutritional value of its meat (Stoyanova et al., 2015; Mahboob et al., 2016; Voigt et al., 2016). In some regions of the planet, such as Brazil, the common carp is considered an invasive species, as it was inadvertently released in the wild and poses a threat to the native aquatic fauna (Smith et al., 2013; Contreras-MacBeath et al., 2014).

Figure 3. Magikarp and Cyprinus carpio.
The shared traits between the Pokémon and the real fish are many: the rounded mouth, the lips, the strong orange color and the presence of barbels (“whiskers”) (Nelson et al., 2016). In China, the common carp is praised as an animal linked to honor and strength, due of its ability to swim against the current; an ancient legend tells about carps that swim upstream, entering through a portal and transforming into dragons (Roberts, 2004). In Pokémon, Magikarp evolves into Gyarados, which resembles a typical Chinese dragon.
Chinchou and Lanturn
Species: Himantolophus sp.; Common name: footballfish.
Chinchou and Lanturn (Fig. 4) were based on fishes of the genus Himantolophus, a group of deep-sea fishes found in almost all oceans living in depths up to 1,800 meters (Klepadlo et al., 2003; Kharin, 2006). These fishes are known as footballfishes, a reference to the shape of their bodies. Fishes of this genus have a special modification on their dorsal fin that displays bioluminescence (the ability to produce light through biological means; Pietsch, 2003), which is used to lure and capture prey (Quigley, 2014). Bioluminescence was the main inspiration for these Pokémon, which have luminous appendages and the Water and Electric types. The sexual dimorphism (difference between males and females) is extreme in these fishes: whilst females reach up to 47 cm of standard-length (that is, body length excluding the caudal fin), males do not even reach 4 cm (Jónsson & Pálsson, 1999; Arronte & Pietsch, 2007).

Figure 4. Chinchou, Lanturn and Himantolophus sp.
Qwilfish
Species: Diodon sp.; Common name: porcupinefish.
Qwilfish (Fig. 5) was based on porcupinefishes, more likely those of the genus Diodon, which present coloring and spines most similar to this Pokémon. Besides the distinctive hard, sharp spines (Fujita et al., 1997), porcupinefishes have the ability to inflate as a strategy to drive off predators (Raymundo & Chiappa, 2000). As another form of defense, these fishes possess a powerful bacterial toxin in their skin and organs (Lucano-Ramírez et al., 2011; Ravi et al., 2016). Accordingly, Qwilfish has both Water and Poison types.

Figure 5. Qwilfish and Diodon sp.
Remoraid
Species: Remora sp.; Common names: remora, suckerfish.
Remoraid was based on a remora (Fig. 6), a fish with a suction disc on its head that allows its adhesion to other animals such as turtles, whales, dolphins, sharks and manta rays (Fertl & Landry, 1999; Silva & Sazima, 2003; Friedman et al., 2013; Nelson et al., 2016). This feature allows the establishment of a commensalisc or mutualisc relationship of transportation, feeding and protection between the adherent species and its “ride” (Williams et al., 2003; Sazima & Grossman, 2006). The similarities also include the name of the Pokémon and the ecological relationship they have with other fish Pokémon: in the same way remoras keep ecological relationships with rays, Remoraid does so with Mantyke and Mantine (Pokémon based on manta rays; see below).

Figure 6. Remoraid and Remora sp.
Mantyke and Mantine
Species: Manta birostris; Common name: manta ray.
The Pokémon Mantyke and its evolved form Mantine (Fig. 7) were probably based on manta rays of the species Manta birostris, which inhabits tropical oceans (Duffy & Abbot, 2003; Dewar et al., 2008) and can reach more than 6 meters of wingspan, being the largest species of ray in existence (Homma et al., 1999; Ari & Correia, 2008; Marshall et al., 2008; Luiz et al., 2009; Nelson et al., 2016). The similarities between the Pokémon and the real fish are: the body shape, the color pattern, the large and distinctive wingspan and even the names.

Figure 7. Mantine, Mantyke and Manta birostris.
Kingdra and Skrelp
Species: Phyllopteryx taeniolatus; Common name: common seadragon.
Kingdra and Skrelp (Fig. 8) were based on the common seadragon. The resemblances between these Pokémon and the real fish species include the leaf-shaped fins that help the animals to camouflage themselves in the kelp “forests” they inhabit (Sanchez-Camara et al., 2006; Rossteuscher et al., 2008; Sanchez-Camara et al., 2011), and the long snout. Also, the secondary type of Kingdra is Dragon. Although both are based on the common seadragon, Kingdra and Skrelp are not in the same “evolutionary line” in the game.
Common seadragons, as the seahorses mentioned above, are of a particular interest to conservationists, because many species are vulnerable due to overfishing, accidental capture and habitat destruction (Foster & Vincent, 2004; Martin-Smith & Vincent, 2006).

Figure 8. Kingdra, Skrelp and Phyllopteryx taeniolatus.
Carvanha
Species: Pygocentrus sp.; Common name: red piranha.
Piranhas of the genus Pygocentrus possibly were the inspiration for the creation of Carvanha (Fig. 9), a Pokémon of voracious and dangerous habits. The main feature shared by the real fish and the Pokémon is the color pattern: bluish in the dorsal and lateral areas, and reddish in the ventral area (Piorski et al., 2005; Luz et al., 2015).
It is worthwhile pointing out that, despite what is shown in movies and other media, piranhas do not immediately devour their prey; instead, they tear off small pieces, bit by bit, such as scales and fins (Trindade & Jucá-Chagas, 2008; Vital et al., 2011; Ferreira et al., 2014).

Figure 9. Carvanha and Pygocentrus sp.
Sharpedo
Order: Carcharhiniformes; Common name: shark.
Sharpedo (Fig. 10), according to its morphological traits (elongated fins), was possibly based on sharks of the order Carcharhiniformes, the largest group of sharks, with 216 species in 8 families and 48 genera. Fishes in this order are common in all oceans, in both coastal and oceanic regions, and from the surface to great depths (Gomes et al., 2010). Several species of Carcharhiniformes are in the IUCN’s (International Union for Conservation of Nature) endangered species list (a.k.a. “Red List”) due to overfishing, as their fins possess high commercial value (Cunningham-Day, 2001).

Figure 10. Sharpedo and a carcharhiniform shark.
Barboach
Species: Misgurnus sp.; Common name: pond loach.
Barboach (Fig. 11) is likely based on fishes of the genus Misgurnus, natively found in East Asia (Nobile et al., 2017) but introduced in several countries (Gomes et al., 2011). These animals, like M. anguillicaudatus Cantor, 1842, are used as ornamental fishes and in folk medicine (Woo Jun et al., 2010; Urquhart & Koetsier, 2014). The shared similarities between the Pokémon and the pond loach include morphological features, such as the elongated body, oval fins and the presence of barbels (Nelson et al., 2016). The resemblance also extends itself to behavior, such as the habit of burying in the mud (Zhou et al., 2009; Kitagawa et al., 2011) and using the barbels to feel the surroundings (Gao et al., 2014). The secondary type of Barboach, Ground, alongside the ability to feel vibrations in the substrate, seem to be a reference to the behavior of the real fishes.

Figure 11. Barboach and Misgurnus sp.
Whiscash
Species: Silurus sp.; Common name: catfish.
Whiscash (Fig. 12) was based on the Japanese mythological creature Namazu, a gigantic catfish that inhabits the underground realm and is capable of creating earthquakes (Ashkenazi, 2003). Namazu also names the Pokémon in the Japanese language (“Namazun”). In Japan, fishes of the genus Silurus are usually associated with this mythological creature and even the common name of these fishes in that country is “namazu” (Yuma et al., 1998; Malek et al., 2004). In addition, the physical traits of the Silurus catfishes also present in Whiscash are the long barbels (or “whiskers”, hence the name Whiscash) and the robust body (Kobayakawa, 1989; Kiyohara & Kitoh, 1994). In addition to the Water type, Whiscash is also Ground type, which is related to Namazu’s fantastic ability of creating earthquakes.

Figure 12. Whiscash and Silurus sp.
Feebas
Species: Micropterus salmoides; Common name: largemouth bass.
The Pokémon Feebas (Fig. 13), a relatively weak fish (as its name implies), was possibly based on a largemouth bass, a freshwater fish native to North America (Hossain et al., 2013). The species was introduced in many countries and is often considered a threat to the native fauna (Welcomme, 1992; Hickley et al., 1994; Godinho et al., 1997; García-Berthou, 2002). Similarities between Feebas and the largemouth bass include the large, wide mouth and the brownish coloration, with darker areas (Brown et al., 2009).

Figure 13. Feebas and Micropterus salmoides.
Milotic
Species: Regalecus sp.; Common name: oarfish.
Often praised as the most beautiful Pokémon of all (Bulbapedia, 2017), Milotic (Fig. 14) certainly lives up to its title. Their long reddish eyebrows were based on the first elongated rays of the dorsal fin of Regalecus species (Nelson et al., 2016), which also share the reddish color of the dorsal fin (Carrasco-Águila et al., 2014). Other similarities between the oarfish and the Pokémon are the elongated body (some oarfishes can grow larger than 3.5 m) and the spots scattered on the body (Chavez et al., 1985; Balart et al., 1999; Dulčić et al., 2009; Ruiz & Gosztonyi, 2010).

Figure 14. Milotic and Regalecus sp.
Huntail
Species: Monognathus sp.; Common name: onejaw.
Probably based on fishes of the genus Monognathus, which have a large mandible and a long dorsal fin (Nelson et al., 2016), Huntail (Fig. 15) is one of the possible evolutionary results of the mollusk Pokémon Clamperl (the other possibility is Gorebyss; see below). According to Raju (1974), fishes of the genus Monognathus live in great depths and have a continuous dorsal fin that ends in an urostyle (“uro” comes from the Greek language and means “tail”, an element also present in the Pokémon’s name).

Figure 15. Huntail and Monognathus sp.
Gorebyss
Family: Nemichthyidae; Common name: snipe eel.
The serpentine body and the thin beak-shaped jaw of Gorebyss (Fig. 16) are features of fishes belonging to the family Nemichthyidae (Nielsen & Smith, 1978). These fishes inhabit tropical and temperate oceans and can be found in depths up to 4,000 meters, in the so-called “abyssal zone” (Cruz-Mena & Anglo, 2016). The Pokémon’s name may be a reference to such habitat.

Figure 16. Gorebyss and a nemichthyid fish.
Relicanth
Species: Latimeria sp.; Common name: coelacanth.
Relicanth (Fig. 17) was based on the coelacanth. The brown coloration, the lighter patches on the body (Benno et al., 2006) and the presence of paired lobed fins (Zardoya & Meyer, 1997) are traits of both the real fish and the Pokémon. It was believed that coelacanths went extinct in the Late Cretaceous, but they were rediscovered in 1938 in the depths off the coast of South Africa (Nikaido et al., 2011). Therefore, the only two living species L. chalumnae Smith, 1939 and L. menadoensis Pouyaud et al., 1999 are known as “living fossils” (Zardoya & Meyer, 1997). Probably for this reason, Relicanth belongs to the Water and Rock types (the “fossil Pokémon” are all Rock-type).

Figure 17. Relicanth and Latimeria sp.
Luvdisc
Species: Helostoma temminckii; Common name: kissing gourami.
The silver-pinkish coloration, the peculiar mouth formed by strong lips and the habit of “kissing” other individuals of their species (which is actually a form of aggression!) are features of the kissing gourami (Sterba 1983; Sousa & Severi 2000; Sulaiman & Daud, 2002; Ferry et al., 2012) that are also seen in Luvdisc (Fig. 18). Helostoma temminckii is native to Thailand, Indonesia, Java, Borneo, Sumatra and the Malay Peninsula (Axelrod et al., 1971), but due to its use an ornamental fish and the irresponsible handling by fishkeepers, it has been introduced in other parts of the world (Magalhães, 2007).

Figure 18. Luvdisc and Helostoma temminckii.
Finneon and Lumineon
Species: Pantodon buchholzi; Common name: freshwater butterflyfish.
Finneon and Lumineon (Fig. 19) were probably based on the freshwater butterflyfish. Finneon has a caudal fin in the shape of a butterfly and Lumineon, like Pantodon buchholzi, has large pectoral fins (Nelson et al., 2016) resembling the wings of a butterfly (hence the popular name of the species). Butterflyfishes are found in West African lakes (Greenwood & Thompson, 1960); their backs are olive-colored while their ventral side is silver, with black spots scattered throughout the body; their fins are pink with some purplish spots (Lévêque & Paugy, 1984). Both Pokémon have color patterns that resemble the freshwater butterflyfish.

Figure 19. Finneon, Lumineon and Pantodon buchholzi.
Basculin
Family: Serrasalmidae; Common name: piranha.
The two forms of the Pokémon Basculin (Fig. 20) seem to have been inspired on fishes from the Serrasalmidae family, such as piranhas. Basculin, like these fishes, has a tall body and conical teeth (Baumgartner et al., 2012). Piranhas are predators with strong jaws that inhabit some South American rivers. Curiously, they are commonly caught by local subsistence fishing (Freeman et al., 2007).

Figure 20. Basculin’s two forms and a serrasalmid fish.
Alomomola
Species: Mola mola; Common name: sunfish.
The very name of this Pokémon is evidence that it was inspired on Mola mola, the sunfish (Fig. 21). Moreover, Alomomola, just like the sunfish, has a circular body with no caudal fin (Pope et al., 2010). The sunfish is the largest and heaviest bony fish in the world, weighting more than 1,500 kg (Freesman & Noakes, 2002; Sims et al., 2009). They inhabit the Atlantic and Pacific Oceans, feeding mainly on zooplankton (Cartamil & Lowe, 2004; Potter & Howell, 2010).
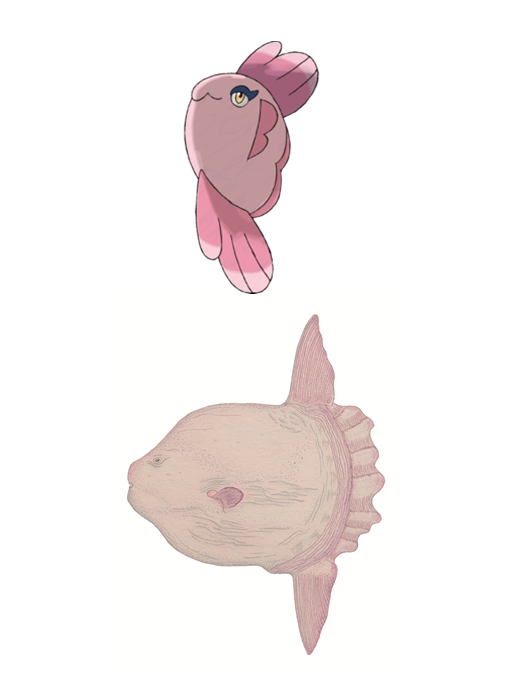
Figure 21. Alomomola and Mola mola.
Tynamo, Eelektrik and Eelektross
Species: Petromyzon marinus; Common name: sea lamprey.
The evolutionary line Tynamo, Eelektrik and Eelektross (Fig. 22) was probably inspired by the life cycle of the sea lamprey, Petromyzon marinus: Tynamo represents a larval stage, Eelektrik a juvenile, and Eelektross an adult. As a larva, the sea lamprey inhabits freshwater environments and, after going through metamorphosis, the juvenile migrates to the ocean, where they start to develop hematophagous (“blood-sucking”) feeding habits (Youson, 1980; Silva et al., 2013). Eelektrik and Eelektross, like the sea lamprey, have a serpentine body and a circular suction cup-mouth with conical teeth. In addition, the yellow circles on the side of these Pokémon resemble the gill slits of lampreys (which are of circular shape) or the marbled spots of P. marinus (Igoe et al., 2004).
It is worth mentioning that Eelektrik and Elektross also seem to possess name and characteristics (Electric type and serpentine body with yellow spots) inspired by the electric eel (Electrophorus electricus Linnaeus, 1766), a fish capable of generating an electrical potential up to 600 volts, making it the greatest producer of bioelectricity in the animal kingdom (Catania, 2014). However, a remarkable characteristic of Eelektrik and Eelektross is the jawless mouth structure of the superclass Petromyzontomorphi species. The electric eel has a jaw and thus belongs to the superclass Gnathostomata (jawed vertebrates) (Gotter et al., 1998).

Figure 22. Tynamo, Eelektrik, Eelektross and P. marinus.
Stunfisk
Order: Pleuronectiformes; Common name: flatfish.
Flattened and predominantly brown in color, Stunfisk (Fig. 23) appears to have been based on fishes of the order Pleuronectiformes. Popularly known as flatfishes, these animals have both eyes on the same side of the head and stay most of their lives buried and camouflaged on sandy and muddy substrates of almost every ocean, feeding on fishes and benthic invertebrates (Sakamoto, 1984; Kramer, 1991; Gibb, 1997). It is likely that the primary type of Stunfisk, Ground, is based on the close relationship between pleuronectiform fishes and the substrate they live in. Species of this group are very valuable for the fishing industry (Cooper & Chapleau, 1998).

Figure 23. Stunfisk and a pleuronectiform fish.
Dragalge
Species: Phycodurus eques; Common name: leafy seadragon.
Dragalge (Fig. 24), a Pokémon belonging to the Poison and Dragon types, was based on a leafy seadragon. This species is found in Australia and it is named after its appearance: this fish has appendages throughout its body that resemble leaves (Larson et al., 2014). This feature, also present in the Pokémon, allows the leafy seadragon to camouflage itself among algae (Wilson & Rouse, 2010). Dragalge is the evolved form of Skrelp, a Pokémon based on a common seadragon (see above).

Figure 24. Dragalge and Phycodurus eques.
Wishiwashi
Species: Sardinops sagax; Common name: Pacific sardine.
Wishiwashi (Fig. 25) was probably based on the Pacific sardine, a pelagic fish with high commercial value and quite abundant along the California and Humboldt Currents (Coleman, 1984; Gutierrez-Estrada et al., 2009; Demer et al., 2012; Zwolinski et al., 2012). The lateral circles of the Pokémon are a reference to the dark spots present on the lateral areas of the real fish (Paul et al., 2001). Furthermore, Wishiwashi has the ability to form a large school, just as sardines do (Emmett et al., 2005; Zwolinski et al., 2007).

Figure 25. Wishiwashi and Sardinops sagax.
Another parallel is the geographic location: the Pokémon belongs to Alola, a fictional region based on Hawaii, and S. sagax is one of the most common sardines in the Eastern Pacific Ocean. From the mid-1920’s to the mid-1940’s, for example, S. sagax supported one of the largest fisheries in the world. The stock collapsed in the late 1940’s, but in the 1990’s it started to recover (McFarlane et al., 2005).
Bruxish
Species: Rhinecanthus rectangulus; Common name: reef triggerfish.
Bruxish (Fig. 26) was probably inspired by the species Rhinecanthus rectangulus, the reef triggerfish of the Hawaiian reefs and other tropical regions (Kuiter & Debelius, 2006; Dornburg et al., 2008). Bruxish has powerful jaws, just like the reef triggerfishes that prey upon a wide variety of invertebrates, such as hard-shelled gastropods, bivalves, echinoderms and crustaceans (Wainwright & Friel, 2000; Froese & Pauly, 2016).

Figure 26. Bruxish and Rhinecanthus rectangulus.
Besides the strong jaw, the overall body shape and the flashy coloring, another parallel can be seen: this Pokémon is an inhabitant of the Alola region (the Pokémon version of Hawaii) and R. rectangulus is actually the state symbol fish of the Hawaiian archipelago (Kelly & Kelly, 1997).
POCKET FISHES UNDER SCRUTINY
The majority of the identified Pokémon (85.29%) is, expectedly, Water-type. A large portion of them (29.41%) was introduced for the first time in the third generation of the franchise, in the Hoenn region.

Figure 27. Representativeness of fish classes in Pokémon.
Only three fish Pokémon were classified in the superclass Petromyzontomorphi (8.82%): the lamprey-like Tynamo, Eelektrik and Eelektross, all of them belonging to the same evolutionary line. In the superclass Gnathostomata, the class Osteichthyes is represented by the highest number of Pokémon: 28 in total (82.35%, Fig. 27). Inside this class, the most representative groups were the order Syngnathiformes (14.71%, Fig. 28), family Syngnathidae (15.63%, Fig. 29) and the genus Petromyzon (10.00%, Fig. 30).

Figure 28. Representativeness of fish orders in Pokémon.
Most of the real fishes on which the Pokémon were based (55.88%, Fig. 31) live in marine environments, followed by freshwater (continental water environments, 32.35%) and finally, brackish water (estuarine environments, 11.76%).
The “fish” species found in the Pokémon world consists of a considerable portion of the ichthyological diversity in our world. According to Nelson et al. (2016), the Osteichthyes class corresponds to 96.1% of all vertebrate fish species (30,508 species), followed by the Condrichthyes with 3.76% (1,197 species) and the Petromyzontida with just 0.14% (46 species). In Pokémon, the proportions of taxa (taxonomic group) that inspired the creatures follow a roughly similar distribution: within the 26 taxa in which the evolutionary families of the Pokémon were based, 23 are Osteichthyes class (88.46%), two are Condrichthyes (7.7%) and one is Petromyzontida (3.84%). If the games follow a pattern of introducing more fish Pokémon over time, it is expected that these proportions will gradually become more equivalent as each new generation of the franchise is released.

Figure 29. Representativeness of fish families in Pokémon.
ALMOST A BIOLOGICAL POCKET-WORLD
Our analysis shows that fish Pokémon are very diverse creatures, both taxonomic and ecologically, despite being a small group within the Pokémon universe (with 801 “species”).
The fish Pokémon are represented by several orders, families and genera of real fishes and, as previously stated, this is actually a relevant sampling of the ichthyological diversity of our planet. The marine Pokémon described here are inhabit from abyssal zones to coastal regions, including reefs. The creative process of the fish monsters in the game must have included a fair share of research on real animals.

Figure 30. Representativeness of fish genera in Pokémon.
The Hoenn region, which has the largest playable surface and includes areas with “too much water”, is also the region with the highest number of fish Pokémon. Furthermore, the majority of these Pokémon live in the marine environment and belongs to the Osteichthyes class, as is observed for real fishes (Nelson et al., 2016; Eschmeyer et al., 2016). However, it is also important to underline that marine fishes are those with the more attractive colors and shapes and, therefore, higher popular appeal, which is vital for a game based in charismatic monsters (Darwall et al., 2011; McClenachan, 2012; Dulvy et al., 2014).

Figure 31. Environments inhabited by the fish Pokémon.
In the present work, the analogy between fish Pokémon and real species allowed a descriptive study of the “Pokéfauna” in a similar manner in which actual faunal surveys are presented. These surveys are an important tool for understanding the structure of communities and to evaluate the conservation status of natural environments (Buckup et al., 2014). It is noteworthy that the association of the monsters with real fishes was only possible because Pokémon have several morphological, ecological and ethological traits that were based on real species.
Pokémon is a successful franchise and many of its staple monsters are already part of the popular imaginary. The creation of the pocket monsters was not done in a random manner; they were mostly inspired by real organisms, particularly animals, and often have specific biological traits taken from their source of inspiration. Thus, analogies between Pokémon and our natural world, such as the ones performed here, open a range of possibilities for science outreach.
REFERENCES
Ari, C. & Correia, J.P. (2008) Role of sensory cues on food searching behavior of a captive Manta birostris (Chondrichtyes, Mobulidae). Zoo Biology 27(4): 294–304.
Arronte, J.C. & Pietsch, T.W. (2007) First record of Himantolophus mauli (Lophiiformes: Himantolophidae) on the slope off Asturias, Central Cantabrian Sea, Eastern North Atlantic Ocean. Cybium 31(1): 85–86.
Ashkenazi, M. (2003) Handbook of Japanese Mythology. ABC-CLIO, Santa Barbara.
Axelrod, H.R.; Emmens, C.W.; Sculthorpe, D.; Einkler, W.V.; Pronek, N. (1971) Exotic Tropical Fishes. TFH Publications, New Jersey.
Balart, E.F.; Castro-Aguirre, J.L.; Amador-Silva, E. (1999) A new record of the oarfish Regalecus kinoi (Lampridiformes: Regalecidae) in the Gulf of California, Mexico. Oceánides 14(2): 137–140.
Baumgartner, G.; Pavanelli, C.S.; Baumgartner, D.; Bifi, A.G.; Debona, T.; Frana, V.A. (2012) Peixes do Baixo Rio Iguaçu: Characiformes. Eduem, Maringá.
Benno, B.; Verheij, E.; Stapley, J.; Rumisha, C.; Ngatunga, B.; Abdallah, A.; Kalombo, H. (2006) Coelacanth (Latimeria chalumnae Smith, 1939) discoveries and conservation in Tanzania. South African Journal of Science 102: 486–490.
Bittencourt, F.; Souza, B.E.; Boscolo, W.E.; Rorato, R.R.; Feiden, A.; Neu, D.H. (2012) Benzocaína e eugenol como anestésicos para o quinguio (Carassius auratus). Arquivo Brasileiro de Medicina Veterinária e Zootecnia 64(6): 1597–1602.
Blackburn, D.G. (1999) Viviparity and oviparity: evolution and reproductive strategies. In: Knobil, E. & Neil, J. D. (Eds.) Encyclopedia of reproduction. Acedemic Press, New York. Pp. 994–1003.
Braga, W.F.; Araújo, J.G.; Martins, G.P.; Oliveira, S.L.; Guimarães, I.G. (2016) Dietary total phosphorus supplementation in goldfish diets. Latin American Journal of Aquatic Research 44(1): 129–136.
Brown, T.G.; Runciman, B.; Pollard, S.; Grant, A.D.A. (2009) Biological synopsis of largemouth bass (Micropterus salmoides). Canadian Manuscript Report of Fisheries and Aquatic Sciences 2884: 1–35.
Buckup, P.A.; Britto, M.R.; Souza-Lima, R.S.; Pascoli, J.C.; Villa-Verde, L.; Ferraro, G.A.; Salgado, F.L.K; Gomes, J.R. (2014) Guia de Identificação das Espécies de Peixes da Bacia do Rio das Pedras, Município de Rio Claro, RJ. The Nature Conservancy, Rio de Janeiro.
Bulbapedia. (2017) Bulbapedia. The community driven Pokémon encyclopedia. Available from: http://bulbapedia.bulbagarden.net/ (Date of access: 20/Jan/2017).
Carrasco-Águila, M.A.; Miranda-Carrillo, O.; Salas-Maldonado, M. (2014) El rey de los arenques Regalecus russelii, segundo ejemplar registrado en Manzanillo, Colima. Ciencia Pesquera 22(2): 85–88.
Cartamil, D.P. & Lowe, C.G. (2004) Diel movement patterns of ocean sunfish Mola mola off southern California. Marine Ecology Progress Series 266: 245–253.
Castro, A.L.C.; Diniz, A.F.; Martins, I.Z.; Vendel, A.L.; Oliveira, T.P.R.; Rosa, I.M.L. (2008) Assessing diet composition of seahorses in the wild using a nondestructive method: Hippocampus reidi (Teleostei: Syngnathidae) as a study-case. Neotropical Ichthyology 6(4): 637–644.
Catania, K. (2014) The shocking predatory strike of the electric eel. Science 346(6214): 1231–1234.
Chávez, H.; Magaña, F.G.; Torres-Villegas, J.R. (1985) Primer registro de Regalecus russelii (Shaw) (Pisces: Regalecidae) de aguas mexicanas. Investigaciones Marinas CICIMAR 2(2): 105–112.
Coleman, N. (1984) Molluscs from the diets of commercially exploited fish off the coast of Victoria, Australia. Journal of the Malacological Society of Australia 6: 143–154.
Contreras-Macbeath, T.; Gaspar-Dillanes, M.T.; Huidobro-Campos, L.; Mejía-Mojica, H. (2014) Peces invasores em el centro de México. In: Mendoza, R. & Koleff, P. (Eds.) Especies Acuáticas Invasoras en México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Ciudad de México. Pp. 413–424.
Cooper, J.A. & Chapleau, F. (1998) Monophyly and intrarelationships of the family Pleuronectidae (Pleuronectiformes), with a revised classification. Fishery Bulletin 96(4): 686–726.
Cruz-Mena, O.I. & Angulo, A. (2016) New records of snipe eels (Anguilliformes: Nemichthyidae) from the Pacific coast of lower Central America. Marine Biodiversity Records 9(1): 1–6.
Cunningham-Day, R. (2001) Sharks in Danger: Global Shark Conservation Status with Reference to Management Plans and Legislation. Universal Plubishers, Parkland.
Darwall, W.R.T.; Holland, R.A.; Smith, K.G.; Allen, D.; Brooks, E.G.E.; Katarya, V.; Pollock, C.M.; Shi, Y.; Clausnitzer, V.; Cumberlidge, N.; Cuttelod, A.; Dijkstra, B.K.; Diop, M.D.; García, N.; Seddon, M.B.; Skelton, P.H.; Snoeks, J.; Tweddle, D.; Vié, J. (2011) Implications of bias in conservation research and investment for freshwater species. Conservation Letters 4: 474–482.
Demer, D.A.; Zwolinski, J.P.; Byers, K.A.; Cutter, G.R.; Renfree, J.S.; Sessions, T.S.; Macewicz, B.J. (2012) Prediction and confirmation of seasonal migration of Pacific sardine (Sardinops sagax) in the California Current Ecosystem. Fishery Bulletin 110(1): 52–70.
Dewar, H.; Mous, P.; Domeler, M.; Muljadi, A.; Pet, J.; Whitty, J. (2008) Movements and site fidelity of the giant manta ray, Manta birostris, in the Komodo Marine Park, Indonesia. Marine Biology 155(2): 121–133.
Dornburg, L.; Santini, F.; Alfaro, M.E. (2008) The influence of model averaging on clade posteriors: an example using the triggerfishes (family Balistidae). Systematic Biology 57(6): 905–919.
Dorward, L.J.; Mittermeier, J.C.; Sandbrook, C.; Spooner, F. (2017) Pokémon GO: benefits, costs, and lessons for the conservation movement. Conservation Letters 10(1): 160–165.
Duffy, C.A.J. & Abbott, D. (2003) Sightings of mobulid rays from northern New Zealand, with confirmation of the occurrence of Manta birostris in New Zealand waters. New Zealand Journal of Marine and Freshwater Research 37(4): 715–721.
Dulčić, J.; Dragičević, B.; Tutman, P. (2009) Record of Regalecus glesne (Regalecidae) from the eastern Adriatic Sea. Cybium 33(4): 339–340.
Dulvy, N.K.; Fowler, S.L.; Musick, J.A.; Cavanagh, R.D.; Kyne, P.M.; Harrison. L.R.; Carlson, J.K.; Davidson. L.N.K.; Fordham, S.V.; Francis, M.P.; Pollock, C.M.; Simpfendorfer, C.A.; Burgess, G.H.; Carpenter, K.E.; Compagno. L.J.V.; Ebert, D.A.; Gibson, C.; Heupel, M.R.; Livingstone, S.R.; Sanciangco. J.C.; Stevens, J.D.; Valenti, S.; White, W.T. (2014) Extinction risk and conservation of the world’s sharks and rays. eLife Sciences 3(e00590): 1–34.
Emmett, R.L.; Blodeur, R.D.; Miller, T.W.; Pool, S.S.; Krutzikowsky, G.K.; Bentley, P.J.; McCrae, J. (2005) Pacific sardine (Sardinops sagax) abundance, distribution, and ecological relationships in the Pacific Northwest. California Cooperative Oceanic Fisheries Investigations Reports 46: 122–143.
Eschmeyer, W.N.; Fricke, R.; van der Laan, R. (2016) Catalog of Fishes: Genera, Species, References. Available from: http://researcharch ive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (Date of access: 25/Nov/ 2016).
Ferreira, F.S.; Vicentin, W.; Costa, F.E.S.; Suárez, Y.R. (2014) Trophic ecology of two piranha species, Pygocentrus nattereri and Serrasalmus marginatus (Characiformes, Characidae), in the floodplain of the Negro River, Pantanal. Acta Limnologica Brasiliensia 26(4): 381–391.
Ferry, L.A.; Konow, N.; Gibb, A.C. (2012) Are kissing gourami specialized for substrate-feeding? Prey capture kinematics of Helostoma temminckii and other anabantoid fishes. Journal of Experimental Zoology 9999A: 1–9.
Fertl, D. & Landry, A.M. Jr. (1999) Sharksucker (Echeneis naucrates) on a bottlenose dolphin (Tursiops truncatus) and a review of other cetacean-remora associations. Marine Mammal Science 15(3): 859–863.
Forsgren, K.L. & Lowe, C.G. (2006) The life history of weedy seadragons, Phyllopteryx taeniolatus (Teleostei: Syngnathidae). Marine and Freshwater Research 57: 313–322.
Foster, S.J. & Vincent, A.C.J. (2004) Life history and ecology of seahorses: implications for conservation and management. Journal of Fish Biology 65(1): 1–61.
Freedman, J.A. & Noakes, D.L.G. (2002) Why are there no really big bony fishes? A point-of-view on maximum body size in teleosts and elasmobranchs. Reviews in Fish Biology and Fisheries 12: 403–416.
Freeman, B.; Nico, L.G.; Osentoski, M.; Jelks, H.L.; Collins, T.M. (2007) Molecular systematics of Serrasalmidae: deciphering the identities of piranha species and unraveling their evolutionary histories. Zootaxa 1484: 1–38.
Friedman, M.; Johanson, Z.; Harrington, R.C.; Near, T.J.; Graham, M.R. (2013) An early fossil remora (Echeneoidea) reveals the evolutionary assembly of the adhesion disc. Proceedings of the Royal Society B 280(1766): 1–8.
Froese, R. & Pauly, D. (2016) FishBase, v. 10/2016. Available from: http://fishbase.org (Date of access: 25/Jan/2017).
Fujita, T.; Hamaura, W.; Takemura, A.; Takano, K. (1997) Histological observations of annual reproductive cycle and tidal spawning rhythm in the female porcupine fish Diodon holocanthus. Fisheries Science 63(5): 715–720.
Gao, L.; Duan, M.; Cheng, F.; Xie, S. (2014) Ontogenetic development in the morphology and behavior of loach (Misgurnus anguillicaudatus) during early life stages. Chinese Journal of Oceanology and Limnology 32(5): 973–981.
García-Berthou, E. (2002) Ontogenetic diet shifts and interrupted piscivory in introduced largemouth bass (Micropterus salmoides). International Review of Hydrobiology 87(4): 353–363.
Gibb, A.C. (1997) Do flatfish feed like other fishes? A comparative study of percomorph prey-capture kinematics. The Journal of Experimental Biology 200: 2841–2859.
Godinho, F.N.; Ferreira, M.T.; Cortes, R.V. (1997) The environmental basis of diet variation in pumpkinseed sunfish, Lepomis gibbosus, and largemouth bass, Micropterus salmoides, along an Iberian river basin. Environmental Biology of Fishes 50(1): 105–115.
Gomes, C.I.D.A.; Peressin, A.; Cetra, M.; Barrela, W. (2011) First adult record of Misgurnus anguillicaudatus Cantor, 1842 from Ribeira de Iguape River Basin, Brazil. Acta Limnologica Brasiliensia 23(3): 229–232.
Gomes, U.L.; Signori, C.N.; Gadig, O.B.F.; Santos, H.R.S. (2010) Guia para Identificação de Tubarões e Raias do Rio de Janeiro. Technical Books Editora, Rio de Janeiro.
Gotter, A.L.; Kaetzel, M.A.; Dedman, J.R. (1998) Electrophorus electricus as a model system for the study of membrane excitability. Comparative Biochemistry and Physiology 119A(1): 225–241.
Greenwood, P.H. & Thompson, K.S. (1960) The pectoral anatomy of Pantodon buchholzi Peters (a freshwater flying fish) and the related Osteoglossidae. Journal of Zoology 135: 283–301.
Gutiérrez-Estrada, J.C.; Yáñez, E.; Pulido-Calvo, I.; Silva, C.; Plaza, F.; Bórquez, C. (2009) Pacific sardine (Sardinops sagax Jenyns, 1842) landings prediction: a neural network ecosystemic approach. Fisheries Research 100: 116–125.
Hickley, P.; North, R.; Muchiri, S.M.; Harper, D.M. (1994) The diet of largemouth bass, Micropterus salmoides, in Lake Naivasha, Kenya. Journal of Fish Biology 44(4): 607–619.
Homma, K.; Maruyama, T.; Itoh, T.; Ishihara, H.; Uchida, S. (1999) Biology of the manta ray, Manta birostris Walbaum, in the Indo-Pacific. In: Séret, B. & Sire, J.-Y. (Eds.) Proceedings of the 5th Indo-Pacific Fish Conference. Ichthyological Society of France, Noumea. Pp. 209–216.
Hossain, M.M.; Perhar, G.; Arhonditsis, G.B.; Matsuishi, T.; Goto, A.; Azuma, M. (2013) Examination of the effects of largemouth bass (Micropterus salmoides) and bluegill (Lepomis macrochirus) on the ecosystem attributes of lake Kawahara-oike, Nagasaki, Japan. Ecological Informatics 18: 149–161.
Igoe, F.; Quigley, D.T.G.; Marnell, F.; Meskell, E.; O’Connor, W.; Byrne, C. (2004) The sea lamprey Petromyzon marinus (L.), river lamprey Lampetra fluviatilis (L.) and brook lamprey Lampetra planeri (Bloch) in Ireland: general biology, ecology, distribution and status with recommendations for conservation. Proceedings of the Royal Irish Academy 104B (3): 43–56.
ITIS. (2016) Integrated Taxonomic Information System. Available from: http://itis.gov/ (Date of access: 25/Nov/2016).
Jónsson, G. & Pálsson, J. (1999) Fishes of the suborder Ceratioidei (Pisces: Lophiiformes) in Icelandic and adjacent waters. Rit Fiskideildar 16: 197–207.
Kasapoglu, N. & Duzgunes, E. (2014) Some population characteristics of long-snouted seahorse (Hippocampus guttulatus Cuvier, 1829) (Actinopterygii: Syngnathidae) in the Southeastern Black Sea. Acta Zoologica Bulgarica 66(1): 127–131.
Kelly, S. & Kelly, T. (1997) Fishes of Hawaii: Coloring Book. Bess Press, Honolulu.
Kent, S.L. (2001) The Ultimate History of Video Games. The Crown Publishing Group, New York.
Kharin, V.E. (2006). Himantolophus sagamius (Himantolophidae), a new fish species for fauna of Russia. Journal of Ichthyology 46(3): 274–275.
Kitagawa, T.; Fujii, Y.; Koizumi, N. (2011) Origin of the two major distinct mtDNA clades of the Japanese population of the oriental weather loach Misgurnus anguillicaudatus (Teleostei: Cobitidae). Folia Zoologica 60(4): 343–349.
Kiyohara, S. & Kitoh, J. (1994) Somatotopic representation of the medullary facial lobe of catfish Silurus asotus as revealed by transganglionic transport of HRP. Fisheries Science 60(4): 393–398.
Klepladlo, C.; Hastings, P.A.; Rosenblatt, R.H. (2003) Pacific footballfish, Himantolophus sagamius (Tanaka) (Teleostei: Himantolophi-dae), found in the surf-zone at Del Mar, San Diego County, California, with notes on its morphology. Bulletin South California Academy of Sciences 102(3): 99–106.
Kobayakawa, M. (1989) Systematic revision of the catfish genus Silurus, with description of a new species from Thailand and Burma. Japanese Journal of Ichthyology 36(2): 155–186.
Kramer, S.H. (1991) The shallow-water flatfishes of San Diego County. California Cooperative Oceanic Fisheries Investigations Reports 32: 128–142.
Kuiter, R.H. & Debelius, H. (2006) World Atlas of Marine Fishes. Hollywood Import and Export, Frankfurt.
Larson, S.; Ramsey, C.; Tinnemore, D.; Amemiya, C. (2014) Novel microsatellite loci variation and population genetics within leafy seadragons, Phycodurus eques. Diversity 6: 33–42.
Lévêque, C. & Paugy, D. (1984) Guide des Poissons d’Eau Douce: de la Zone du Programme de Lutte contre l’Onchocercose em Afrique de l’Ouest. ORSTOM, Paris.
Lucano-Ramírez, G.; Peña-Pérez, E.; Ruiz-Ramírez, S.; Rojo-Vázquez, J.; González-Sansón, G. (2011) Reproducción del pez erizo, Diodon holocanthus (Pisces: Diodontidae) en la plataforma continental del Pacífico Central Mexicano. Revista de Biologia Tropical 59 (1): 217–232.
Luiz, O.J. Jr.; Balboni, A.P.; Kodja, G.; Andrade, M.; Marum, H. (2009) Seasonal occurrences of Manta birostris (Chondrichthyes: Mobulidae) in southeastern Brazil. Ichthyological Research 56(1): 96–99.
Luz, L.A.; Reis, L.L.; Sampaio, I.; Barros, M.C.; Fraga, E. (2015) Genetic differentiation in the populations of red piranha, Pygocentrus nattereri Kner (1860) (Characiformes: Serrasalminae), from the river basins of northeastern Brazil. Brazilian Journal of Biology 75(4): 838–845.
Magalhães, A.L.B. (2007) Novos registros de peixes exóticos para o Estado de Minas Gerais, Brasil. Revista Brasileira de Zoologia 24(1): 250–252.
Mahboob, S.; Kausar, S.; Jabeen, F.; Sultana, S.; Sultana, T.; Al-Ghanin, K.A.; Hussain, B.; Al-Misned, F.; Ahmed, Z. (2016) Effect of heavy metals on liver, kidney, gills and muscles of Cyprinus carpio and Wallago attu inhabited in the Indus. Brazilian Archives of Biology and Technology 59(e16150275): 1–10.
Malek, M.A.; Nakahara, M.; Nakamura, R. (2004) Uptake, retention and organ/tissue distribution of 137Cs by Japanese catfish (Silurus asotus Linnaeus). Journal of Environmental Radioactivity 77(2): 191–204.
Marshall, A.D.; Pierce, S.J.; Bennett, M.B. (2008) Morphological measurements of manta rays (Manta birostris) with a description of a foetus from the east coast of Southern Africa. Zootaxa 1717: 24–30.
Martin-Smith, K.M. & Vincent, A.C.J. (2006) Exploitation and trade of Australian seahorses, pipehorses, sea dragons and pipefishes (family Syngnathidae). Oryx 40(2): 141–151.
McClenachan, L.; Cooper, A.B.; Carpenter, K.E.; Dulvy, N.K. (2012) Extinction risk and bottlenecks in the conservation of charismatic marine species. Conservation Letters 5: 73–80.
McFarlane, G.A.; MacDougall, L.; Schweigert, J.; Hrabok, C. (2005) Distribution and biology of Pacific sardines (Sardinops sagax) off British Columbia, Canada. California Cooperative Oceanic Fisheries Investigations 46: 144–160.
Moreira, R.L.; da Costa, J.M.; Teixeira, E.G.; Moreira, A.G.L.; De Moura, P.S.; Rocha, R.S.; Vieira, R. H.S.F. (2011) Performance of Carassius auratus with diferent food strategies in water recirculation system. Archivos de Zootecnia 60(232): 1203–1212.
Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. (2016) Fishes of the World. Wiley, New Jersey.
Nielsen, J.G. & Smith, D.G. (1978) The eel family Nemichthyidae (Pisces, Anguilliformes). Dana Report 88: 1–71.
Nikaido, M.; Sasaki, T.; Emerson, J.J.; Aibara, M.; Mzighani, S.I.; Budeba, Y.L.; Ngatunga, B.P.; Iwata, M.; Abe, Y.; Li, W.H.; Okada, N. (2011) Genetically distinct coelacanth population off the northern Tanzanian coast. Proceedings of the National Academy of Sciences of the United States 108(44): 18009–18013.
Nobile, A.B.; Freitas-Souza, D.; Lima, F.P.; Bayona Perez, I.L.; Britto, S.G.C.; Ramos, I.P. (2017) Occurrence of Misgurnus anguillicaudatus (Cantor, 1842) (Cobitidae) in the Taquari River, upper Paraná Basin, Brazil. Journal of Applied Ichthyology (in press).
Official Pokémon Website, The. (2016) The Official Pokémon Website. Available from: http://poke mon.com/ (Date of access: 20/Nov/2016).
Ortega-Salas, A.A. & Reyes-Bustamante, H. (2006) Initial sexual maturity and fecundity of the goldfish Carassius auratus (Perciformes: Cyprynidae) under semi-controlled conditions. Revista de Biologia Tropical 54(4): 1113–1116.
Paul, L.J.; Taylor, P.R.; Parkinson, D.M. (2001) Pilchard (Sarditlops neopilchardus) biology and fisheries in New Zealand, and a review of pilchard (Sardinops, Sardina) biology, fisheries, and research in the main world fisheries. New Zealand Fisheries Assessment Report 37: 1–44.
Pietsch, T.W. (2003) Himantolophidae. Footballfishes (deepsea anglerfishes). In: Carpenter, K.E. (Ed.) FAO Species Identification Guide for Fishery Purposes. The Living Marine Resources of The Western Central Atlantic. Vol. 2: Bony Fishes Part 1 (Acipenseridae to Grammatidae). Food and Agriculture Organization of the United Nations, Rome. Pp. 1060–1061.
Piorski, N.M.; Alves, J.L.R.; Machado, M.R.B.; Correia, M.M.F. (2005) Alimentação e ecomorfologia de duas espécies de piranhas (Characiformes: Characidae) do lago de Viana, estado do Maranhão, Brasil. Acta Amazonica 35(1): 63–70.
Pope, E.C.; Hays, G.C.; Thys, T.M.; Doyle, T.K.; Sims, D.S.; Queiroz, N.; Hobson, V.J.; Kubicek, L.; Houghton, J.D.R. (2010) The biology and ecology of the ocean sunfish Mola mola: a review of current knowledge and future research perspectives. Reviews in Fish Biology and Fisheries 20(4): 471–487.
Potter, I.F. & Howell, W.H. (2010) Vertical movement and behavior of the ocean sunfish, Mola mola, in the northwest Atlantic. Journal of Experimental Marine Biology and Ecology 396(2): 138–146.
Quigley, D.T. (2014) Ceratioid anglerfishes (Lophiiformes: Ceratioidei) in Irish waters. Sherkin Comment 58: 1–7.
Raju, S.N. (1974) Three new species of the genus Monognathus and the Leptocephali of the order Saccopharyngiformes. Fishery Bulletin 72(2): 547–562.
Ravi, L.; Manu, A.; Chocalingum, R.; Menta, V.; Kumar, V.; Khanna, G. (2016) Genotoxicity of tetrodotoxin extracted from different organs of Diodon hystrix puffer fish from South East Indian Coast. Research Journal of Toxins 8(1): 8–14.
Raymundo, A.R. & Chiappa, X. (2000) Hábitos alimentarios de Diodon histrix y Diodon holocanthus (Pisces: Diodontidae) en las costas de Jalisco y Colima, México. Boletín del Centro de Investigaciones Biológicas 34(2): 181–210.
Roberts, J. (2004) Chinese Mythology A to Z. Facts on File, New York.
Rosa, I.L.; Oliveira, T.P.R; Castro, A.L.C.; Moraes, L.E.S.; Xavier, J.H.A.; Nottingham, M.C.; Dias, T.L.P.; Bruto-Costa, L.V.; Araújo, M.E.; Birolo, A.B; Mai, A.C.G; Monteiro-Neto, C. (2007) Population characteristics, space use and habitat associations of the seahorse Hippocampus reidi (Teleostei: Syngnathidae). Neotropical Icthyology 5(3): 405–414.
Rosa, I.L.; Sampaio, C.L.S.; Barros, A.T. (2006) Collaborative monitoring of the ornamental trade of seahorses and pipefishes (Teleostei: Syngnathidae) in Brazil: Bahia state as a case study. Neotropical Icthyology 4(2): 247–252.
Rossteucher, S.; Wenker, C.; Jermann, T.; Wahli, T.; Oldenberg, E.; Schmidt-Posthaus, H. (2008) Severe scuticociliate (Philasterides dicentrarchi) infection in a population of sea dragons (Phycodurus eques and Phyllopteryx taeniolatus). Veterinary Pathology 45(4): 546–550.
Ruiz, A.E. & Gosztonyi, A.E. (2010) Records of regalecid fishes in Argentine Waters. Zootaxa 2509: 62–66.
Sakamoto, K. (1984) Interrelationships of the family Pleuronectidae (Pisces: Pleuronectiformes). Memoirs of Faculty of Fisheries of Hokkaido University 31(1/2): 95–215.
Sanchez-Camara, J. & Booth, D.J. (2004) Movement, home range and site fidelity of the weedy seadragon Phyllopteryx taeniolatus (Teleostei: Syngnathidae). Environmental Biology of Fishes 70(1): 31–41.
Sanchez-Camara, J.; Booth, D.J.; Murdoch, J.; Watts, D.; Turon, X. (2006) Density, habitat use and behaviour of the weedy seadragon Phyllopteryx taeniolatus (Teleostei: Syngnathidae) around Sydney, New South Wales, Australia. Marine and Freshwater Research 57: 737–745.
Sanchez-Camara, J.; Booth, D.J.; Turon, X. (2005) Reproductive cycle and growth of Phyllopteryx taeniolatus. Journal of Fish Biology 67(1): 133–148.
Sanchez-Camara, J.; Martin-Smith, K.; Booth, D.J.; Fritschi, J.; Turon, X. (2011) Demographics and vulnerability of a unique Australian fish, the weedy seadragon Phyllopteryx taeniolatus. Marine Ecology Progress Series 422: 253–264.
Sazima, I. & Grossman, A. (2006) Turtle riders: remoras on marine turtles in Southwest Atlantic. Neotropical Ichthyology 4(1): 123–126.
Schlesinger, H. (1999a) Pokémon Fever: The Unauthorized Guide. St. Martin’s Paperbacks, New York.
Schlesinger, H. (1999b) How to Become a Pokémon Master. St. Martin’s Paperbacks, New York.
Silva, S.; Servia, M.J.; Vieira-Lanero, R.; Barca, S.; Cobo, F. (2013) Life cycle of the sea lamprey Petromyzon marinus: duration of and growth in the marine life stage. Aquatic Biology 18: 59–62.
Silva-Jr., J.M. & Sazima, I. (2003) Whalesuckers and a spinner dolphin bonded for weeks: does host fidelity pay off? Biota Neotropica 3(2): 1–5.
Sims, D.W.; Queiroz, N.; Doyle, T.K.; Houghton, J.D.R.; Hays, G.C. (2009) Satellite tracking of the world’s largest bony fish, the ocean sunfish (Mola mola L.) in the North East Atlantic. Journal of Experimental Marine Biology and Ecology 370: 127–133.
Smith, W.S.; Biagioni, R.C.; Halcsik, L. (2013) Fish fauna of Floresta Nacional de Ipanema, São Paulo State, Brazil. Biota Neotropica 13(2): 175–181.
Soares, C.M.; Hayashi, C.; Gonçalves, G.S.; Galdioli, E.M.; Boscolo, W.R. (2000) Plâncton, Artemia sp., dieta artificial e suas combinações no desenvolvimento e sobrevivência do quinguio (Carassius auratus) durante a larvicultura. Acta Scientiarum 22(2): 383–388.
Sousa, W.T.Z. & Severi, W. (2000) Desenvolvimento larval inicial de Helostoma temminckii Cuvier & Valenciennes (Helostomatidae, Perciformes). Revista Brasileira de Zoologia 17(3): 637–644.
Sterba, G. (1983) The Aquarium Encyclopedia. MIT Press, Cambridge.
Stoyanova, S.; Yancheva, V.S.; Velcheva, I.; Uchikova, E.; Georgieva, E. (2015) Histological alterations in common carp (Cyprinus carpio Linnaeus, 1758) gills as potential biomarkers for fungicide contamination. Brazilian Archives of Biology and Technology 58(5): 757–764.
Sulaiman, Z.H. & Daud, H.K.H. (2002) Pond aquaculture of kissing gouramis Helostoma temminckii (Pisces: Helostomatidae) in Bukit Udal, Tutong: a preliminary investigation. Bruneiana 3: 34–41.
Tobin, J. (2004) Pikachu’s Global Adventure: The Rise and Fall of Pokémon. Duke University Press, Durham.
Trindade, M.E.J. & Jucá-Chagas, R. (2008) Diet of two serrasalmin species, Pygocentrus piraya and Serrasalmus brandtii (Teleostei: Characidae), along a stretch of the Rio de Contas, Bahia, Brazil. Neotropical Ichthyology 6(4): 645–650.
Urquhart, A.N. & Koetsier, P. (2014) Diet of a cryptic but widespread invader, the oriental weatherfish (Misgurnus anguillicaudatus) in Idaho, USA. Western North American Naturalist 74(1): 92–98.
Vital, J.F.; Varella, A.M.B.; Porto, D.B.; Malta, J.C.O. (2011) Sazonalidade da fauna de metazoários de Pygocentrus nattereri (Kner, 1858) no Lago Piranha (Amazonas, Brasil) e a avaliação de seu potencial como indicadora da saúde do ambiente. Biota Neotropica 11(1): 199–204.
Voigt, C.L.; Silva, C.P.; Campos, S.X. (2016) Avaliação da bioacumulação de metais em Cyprinus carpio pela interação com sedimento e água de reservatório. Química Nova 39(2): 180–188.
Wainwright, P.C. & Friel, J.P. (2000) Effects of prey type on motor pattern variance in tetraodontiform fishes. Journal of Experimental Zoology 286(6): 563–571.
Welcomme, R.L. (1992) A history of international introductions of inland aquatic species. ICES Marine Science Symposia 194: 3–14.
Whitehill, S.; Neves, L.; Fang, K.; Silvestri, C. (2016) Pokémon: Visual Companion. The Pokémon Company International / Dorling Kindersley, London.
Williams, E.H.; Mignucci-Giannoni, A.A.; Bunkley-Williams, L.; Bonde, R.K.; Self-Sullivan, C.; Preen, A.; Cockcroft, V.G. (2003) Echeneid-sirenian associations, with information on sharksucker diet. Journal of Fish Biology 63(5): 1176–1183.
Wilson, N.G. & Rouse, G.W. (2010) Convergent camouflage and the non-monophyly of ‘seadragons’ (Syngnathidae: Teleostei): suggestions for a revised taxonomy of syngnathids. Zoologica Scripta 39(6): 551–558.
Woo Jun, J.; Hyung Kim, J. Gomez, D.K.; Choresca, C.H. Jr.; Eun Han, J.; Phil Shin, S.; Chang Park, C. (2010) Occurrence of tetracycline-resistant Aeromonas hydrophila infection in Korean cyprinid loach (Misgurnus anguillicaudatus). African Journal of Microbiology Research 4(9): 849–855.
Yuma, M.; Hosoya, K.; Nagata, Y. (1998) Distribution of the freshwater fishes of Japan: an historical review. Environmental Biology of Fishes 52(1): 97–124.
Zardoya, R. & Meyer, A. (1997) The complete DNA sequence of the mitochondrial genome of a “living fossil,” the coelacanth (Latimeria chalumnae). Genetics 146: 995–1010.
Zhou, X.; Li, M.; Abbas, K.; Wang, W. (2009) Comparison of haematology and serum biochemistry of cultured and wild dojo loach Misgurnus anguillicaudatus. Fish Physiology and Biochemistry 35(3): 435–441.
Zwolinski J.P.; Demer, D.A.; Byers, K.A.; Cutter, G.R.; Renfree, J.S.; Sessions, T.S.; Macewicz, B.J. (2012) Distributions and abundances of Pacific sardine (Sardinops sagax) and other pelagic fishes in the California Current Ecosystem during spring 2006, 2008, and 2010, estimated from acoustic-trawl surveys. Fishery Bulletin 110(1): 110–122.
Zwolinski, J.P.; Morais, A.; Marques, V.; Stratoudakis, Y.; Fernandes, P.G. (2007) Diel variation in the vertical distribution and schooling behaviour of sardine (Sardina pilchardus) off Portugal. Journal of Marine Science 64(5): 963–972.
FURTHER READING
Balmford, A.; Clegg, L.; Coulson, T.; Taylor, J. (2002) Why conservationists should heed Pokémon. Science 295: 2367.
Shelomi, M.; Richards, A.; Li, I.; Okido, Y. (2012) A phylogeny and evolutionary history of the Pokémon. Annals of Improbable Research 18(4): 15–17.
ABOUT THE AUTHORS
Augusto Mendes began his journey as a Pokémon trainer in his childhood, when his parents gave him a green Game Boy Color with Pokémon Red for Christmas. Currently, he is a master’s degree student in the Program of Marine Biology and Coastal Environments of UFF, where he works with zooarchaeology of fishes and education.
Felipe Guimarães is in love with Pokémon (since he first watched the TV series) and the natural world. He graduated in Biology from the UERJ, where he worked with taxonomy and ecology of fishes. He also works with popularization of science and environmental education.
Clara Eirado-Silva, when she was eight years old, told her parents she would study sharks. She has always been passionate about art too and draw since her childhood. Currently, she holds a “Junior Science” scholarship, working on fishing ecology with emphasis on reproductive biology. In her free time, she draws her much loved fishes.
Although Pokémon is not exactly Dr. Edson Silva’s cup of tea, he watched all movies with his daughter, who’s crazy about the little monsters. As fate would have it, his work on population genetics of marine organisms attracted a master’s student (A.B.M.) who’s an equally crazy pokéfan. May Arceus not spare him from the monsters!