Henry N. Thomas
University of California, Berkeley, USA.
Email: h.thomas (at) berkeley (dot) edu
https://doi.org/10.5281/zenodo.8241615
Rodan is a giant monster, or kaiju, created by Toho Studios (Tokyo, Japan). First appearing in 1956 in its own movie (aptly named Rodan), the monster has since become strongly associated with the Godzilla franchise (Fig. 1). Rodan has appeared in seven movies (not counting cameos through stock footage), including the recently released Godzilla: King of the Monsters (2019).

Although often compared to birds, Rodan is explicitly based on pterosaurs. In fact, its Japanese name ラドン Radon is a contraction of the name of the pterosaur genus Pteranodon, and its design reflects this. Rodan’s head, with its toothless beak and curved crests, is clearly based on that of Pteranodon. The original Rodan’s crest shape bears a particular resemblance to specimen YPM 2594[1], a fossil of an adult male Pteranodon longiceps (Fig. 2).
COMPARISON TO ACTUAL PTEROSAURS
Anatomy
Of course, given the first movie starring Rodan came out in 1956, when Pterosaurology was still in its infancy, it is by no means an accurate depiction of a pterosaur. The first difference, obviously, is size. Depending on the incarnation, Rodan has a wingspan between 120 and 265 meters (Table 1 and Fig. 3; Wikizilla, 2019). The very largest pterosaurs belonged to family Azhdarchidae, with giants such as Quetzalcoatlus and Arambourgiania (Fig. 2) reaching roughly 11-meter wingspans. This is near the largest estimated size a pterosaur can get and be able to launch itself off the ground and stay airborne (Witton & Habib, 2010). Rodan is also very different proportionally from the largest pterosaurs known. Azhdarchids generally had giant heads, long necks, very long limbs, and tiny, tiny bodies (Habib, 2019). Rodan, in contrast, has relatively short limbs and a long torso. Given from 1956 to 2004 he was portrayed by the “suitmation” typical of tokusatsu movies (Fig. 1), this is understandable from an out-of-universe perspective.



Rodan’s skull is much smaller proportionally than most pterosaurs. Even small-headed pterosaurs have large skulls proportionally, compared to other reptiles. In particular, large pterosaurs like azhdarchids, anhanguerids, Pteranodon (Fig. 4), and Thalassodromeus have proportionally massive skulls. Huge heads, surprisingly, conveyed certain advantages. They likely made takeoff easier – pterosaurs took off by catapulting themselves into the air from a quadrupedal position, and large heads would make this easier by shifting the center of gravity forwards. In flight, the massive heads were relatively light and aerodynamic (Habib, 2019). As Rodan takes off bipedally – a consequence of being portrayed by suitmation for 48 years – the same benefits may not apply.

Rodan’s skull also has both a toothless anterior beak and posterior teeth. Pteranodon does not have teeth (its name literally means “toothless wing”), and neither do azhdarchids (Fig. 2). A combination of a toothless anterior beak and posterior teeth is only known in two pterosaur families: Dsungaripteridae and Rhamphorhynchidae, both of which are much smaller. Like many pterosaurs, including Pteranodon, Rodan also has a crest on the posterior end of its skull. However, in all pterosaurs with posterior head crests, there is only one crest, projecting from the midline of the skull. This crest is paired in most of Rodan’s appearances. The sole exception is in Godzilla vs. Mechagodzilla II, where Rodan has three crests: one midline crest and two smaller crests flanking it. Presumably, this was done because it looks cool, and not as a bid for scientific accuracy.
Rodan walks upright on two legs in almost every appearance (Fig. 1), a consequence of being portrayed via suitmation. This was accurate at one point (e.g., Padian, 1983), but trackways and biomechanics now show that all pterosaurs walked quadrupedally. That said, Rodan always stands plantigrade, as real pterosaurs did (Witton, 2015). Surprisingly, though, in its original appearance, Rodan’s wing membranes appear to go all the way down to the ankles. This is in line with fossil evidence, which shows that pterosaur wing membranes attached at the ankles instead of the hips or knees (Elgin et al., 2011). The wing membranes are also portrayed as thick and fleshy, which is similar to the wing membranes (patagia) of actual pterosaurs. Pterosaur patagia were more complex than the thin, veiny wings they’re sometimes portrayed with – they had a muscular layer and were supported by fibers called aktinofibrils (Witton, 2013). These two traits of the wing membrane are things even recent movies like Jurassic World (2015) get wrong, and it’s likely Toho got them right by accident.
In almost every appearance, Rodan is covered by bare, leathery skin, with large pointed plates (scutes) on the belly. This is in contrast to actual pterosaurs, which as far as we know were universally covered in a type of fuzz called pycnofibres. This fuzz likely shares an evolutionary origin with the feathers of birds, albeit not being as complex (Yang et al., 2019). Large scutes would add weight and probably negatively impact flight ability. However, the recent Godzilla: King of the Monsters takes a new and interesting direction, where Rodan is instead covered with a rocky armor. It is implied in supplementary marketing material that this armor is tied to a magmatic physiology, which we can probably safely say was not present in any real pterosaur.
As a side note, there is a lone outlier in portrayals of Rodan. The opening montage of Godzilla: Planet of the Monsters (2017) briefly shows the skeleton of Rodan, and it is identical to that of a male Pteranodon longiceps. Presumably, it would have also been identical in life.
Behavior
As a giant monster, Rodan’s behavior does not accurately reflect that of real pterosaurs either. Rodan can create powerful gusts of wind and destructive shockwaves in flight, which would have been impossible for any real pterosaur (Mike Habib, pers. comm.). In Godzilla vs. Mechagodzilla II (1993), when exposed to radiation, Rodan turns into Fire Rodan and gains a radioactive heat beam ability, similar to Godzilla’s famous atomic fire breath. A weapon like this may be analogous to the defensive acid fired from the abdomen of the bombardier beetle; both are superheated and explosively fired (Dean et al., 1990). Although the presence of radioactive energy weapons does not preserve in the fossil record, it would be unprecedented within vertebrates, and we can probably safely presume no real pterosaurs were armed with one.
The original Rodan portrays two individual Rodans, a male and a female. The two are only different in coloration; one is a richer burgundy than the other. Drastic sexual dimorphism is known in some pterosaurs, such as Pteranodon and Hamipterus, where the crests and overall body size of the males were significantly larger than those of the females (Bennett, 1992; Wang et al., 2014). Other pterosaur groups, such as azhdarchoids, appear to show no such skeletal dimorphism (Manzig et al., 2014; Brian Andres, pers. comm.), but this does not rule out other forms of sexual dimorphism, such as differing colorations. The two Rodans are implied to mate for life; when one falls into a volcano, the other sacrifices itself to join its mate in death. It is unknown whether any pterosaurs mated for life. The structure of Pteranodon populations, with more female individuals than males, implies polygamous lek mating similar to sea lions or grouse (Bennett, 1992), while the even sex makeup of Hamipterus colonies implies they may have been monogamous like modern seabirds (Wang et al., 2014).
There is one thing Toho got right with Rodan’s behavior, however. In Rodan, an egg is discovered in a mine in Kitamatsu, on the southern Japanese island of Kyushu. The egg hatches, and the hatchling Rodan already has fully-developed wings and the ability to hunt Meganulons (a fictional species of giant prehistoric insects) on its own. Numerous fossils of pterosaur babies – also called flaplings – have been discovered. These flaplings already show adaptations for full flight, and it is thus likely that pterosaurs were superprecocial, i.e., babies could fend for themselves immediately after hatching (Witton, 2013). Toho seems to have gotten superprecocial pterosaurs right by accident, given this wasn’t recognized until decades later. It’s almost surprising; given Rodan’s behavior generally takes cue from modern birds, one might have expected Rodan flaplings to be altricial, like birds of prey. As well, Destroy All Monsters (1968) portrays Rodan feeding on cetaceans, presumably an upgrade from the piscivory of large oceanic pterosaurs such as Pteranodon (Bennett, 1994). In contrast, large terrestrial pterosaurs such as azhdarchids were likely omnivorous or carnivorous, foraging for food on the ground instead of taking it from the air (Witton & Naish, 2008).
SPECULATIVE EVOLUTION
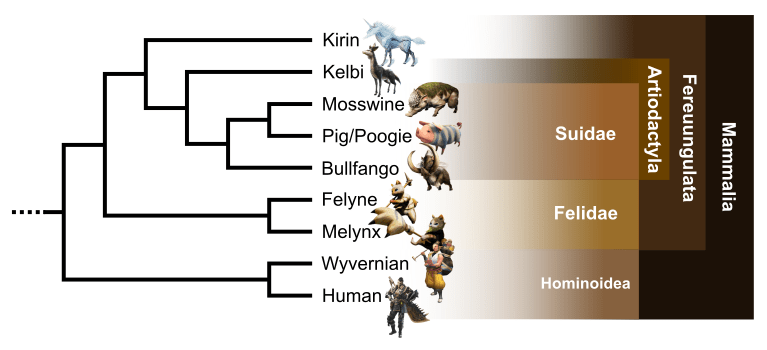
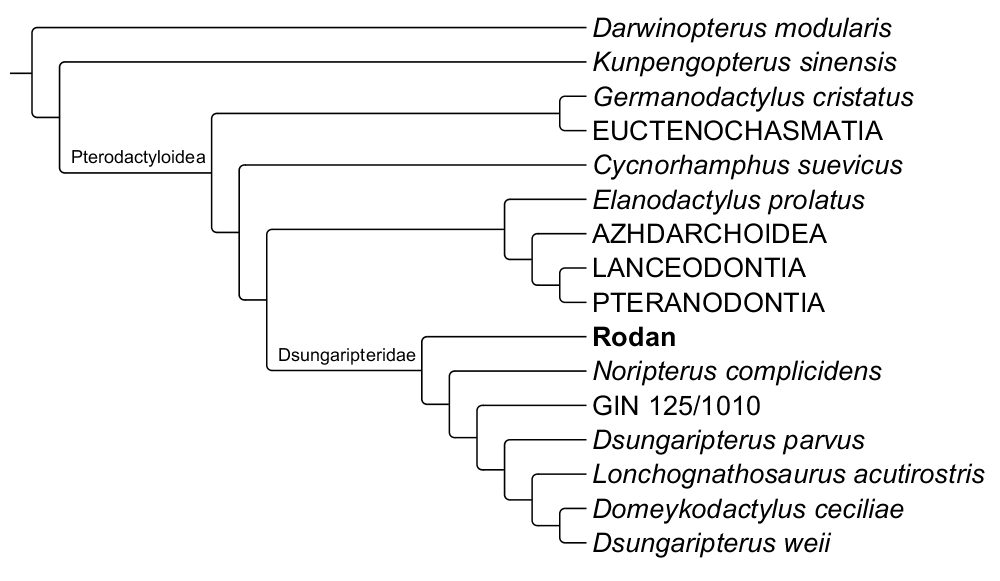
Suppose, for a second, Rodan was real. If you’re anything like me, you may ask: where would it fall on the pterosaur family tree? To answer this question, I coded Rodan into a phylogenetic analysis of pterosaurs I’ve been working on (Thomas, 2018), using what anatomy I could infer from its external appearance. Rodan ended up nowhere near Pteranodon or the giant azhdarchids, but inside Dsungaripteridae (Fig. 5). Dsungaripterids are the only pterodactyloids that have both toothless anterior beaks and posterior teeth, as Rodan does. The subtriangular skull outline and posterior crest of Dsungaripterus (Fig. 2) and Noripterus compare well with the outline of Rodan’s head. Dsungaripterids are also noteworthy for having unusually thickened bone walls (at least 3 mm, compared to less than 2 mm in most pterosaurs) and overall being more robust than most pterodactyloids, traits that would befit a kaiju-sized pterosaur.
How might a real pterosaur resembling Rodan evolve? Let us speculate for a moment. We’ll start with a dsungaripterid – perhaps the proportionally smallest-headed of all pterodactyloids. Increase the size, as well as the proportional wingspan. We can make a few changes to the shape of the skull, including emphasizing the brow ridge and pairing the crests. Given pterosaurs were covered with pycnofibres, and scutes of any sort would weigh down the animal, it would be unlikely to evolve in a pterosaur. We can emulate Rodan’s chest scutes with stiff bristles, however; similar stiffened filaments can be found in other avemetatarsalians, such as a few bird species and the ornithschian Psittacosaurus (Mayr et al., 2016). We can also give this pterosaur stronger hindlimbs, and the habit of rearing bipedally, to emulate the classic suitmation portrayal of Rodan (Fig. 6).

PUBLIC RELATIONS
Since its first appearance, Rodan has become one of the more popular and recognizable giant monsters. It is considered one of the “big five” Toho kaiju, alongside fellow superstars Godzilla, Mothra, Mechagodzilla, and King Ghidorah. But is the portrayal of Rodan a cause for concern among science communicators? Probably not. We’re talking about a pterosaur with a wingspan in the hundreds of meters who creates hurricane-force winds and can sometimes breathe atomic energy. The laws of physics are gleefully trodden upon in Toho’s kaiju movies. It’s fair to assume that nobody expects accurate scientific information from them, and from 1960 onwards the movies themselves never act like this is the case. It’s as if Toho knows its movies are not the best place for science. A greater issue, I would say, would be media that purport to show real species of pterosaur, and yet propagate and enable common misconceptions anyways.
REFERENCES
Bennett, S.C. (1992) Sexual dimorphism of Pteranodon and other pterosaurs, with comments on cranial crests. Journal of Vertebrate Palentology 12(4): 422–434.
Bennett, S.C. (1994) The pterosaurs of the Niobrara Chalk. The Earth Scientist 11(1): 22–25.
Dean, J.; Aneshansley, D.J.; Edgerton, H.E.; Eisner, T. (1990) Defensive spray of the bombardier beetle: a biological pulse jet. Science 248(4960): 1219–1221.
Elgin, R.A.; Hone, D.W.E.; Frey, E. (2011) The extent of the pterosaur flight membrane. Acta Palaeontologica Polonica 56(1): 99–111.
Habib, M.P. (2019) Giant flying jaws: aerodynamic effects and constraints on cranial hypertrophy in pterosaurs. 11th North American Paleontological Convention, Riverside.
Manzig, P.C.; Kellner, A.W.A.; Weinschutz, L.C.; Fragoso, C.E.; Vega, C.S.; Guimaraes, G.B.; Godoy, L.C.; Liccardo, A.; Ricetti, J.H.Z.; de Moura, C.C. (2014) Discovery of a rare pterosaur bone bed in a Cretaceous desert with insights on ontogeny and behavior of flying reptiles. PLoS ONE 9(8): e100005.
Mayr, G.; Pittman, M.; Saitta, E.; Kaye, T.G.; Vinther, J. (2016) Structure and homology of Psittacosaurus tail bristles. Palaeontology 59(6): 793–802.
Padian, K. (1983) A functional analysis of flying and walking in pterosaurs. Paleobiology 9(3): 218–239.
Thomas, H.N. (2018) A novel phylogenetic analysis of azhdarchoid pterosaurs, with comments on their biogeography and paleoecology. 1st Palaeontological Virtual Conference, Valencia.
Wang, X.; Kellner, A.W.A.; Jiang, S.; Wang, Q.; Ma, X.; Paidoula, Y.; Cheng, X.; Rodrigues, T.; Meng, X.; Zhang, J.; Li, N.; Zhou, Z. (2014) Sexually dimorphic tridimensionally preserved pterosaurs and their eggs from China. Current Biology 24(12): 1323–1330.
Wikizilla. (2019) Rodan. Wikizilla, the kaiju encyclopedia. Available from: https://wikizilla.org/wiki/Rodan (Date of access: 31/Aug/2019).
Witton, M.P. (2013) Pterosaurs: Natural History, Evolution, Anatomy. Princeton University Press, Princeton.
Witton, M.P. (2015) Were early pterosaurs inept terrestrial locomotors? PeerJ 3: e1018.
Witton, M.P. & Habib, M.B. (2010) On the size and flight diversity of giant pterosaurs, the use of birds as pterosaur analogues and comments on pterosaur flightlessness. PLoS ONE 5(11): e13982.
Witton, M.P. & Naish, D. (2008) A reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS ONE 3(5): e2271.
Yang, Z.; Jiang, B.; McNamara, M.E.; Kearns, S.L.; Pittman, M.; Kaye, T.G.; Orr, P.J.; Xu, X.; Benton, M.J. (2019) Pterosaur integumentary structures with complex feather-like branching. Nature Ecology & Evolution 3: 24–30.
Acknowledgements
I would like to thank Charlotte Bowman, Luigi Gaskell, Scott Reid and Tristan Stock for proofreading the manuscript and offering feedback, and again Scott Reid for the artwork.
About the author
Henry Thomas is a biology student at the University of California, Berkeley, and the world’s leading expert on Rodan phylogeny. His main research interest is pterosaurs.
[1] The specimen is kept in the collection of Yale Peabody Museum of Natural History (New Haven, USA).