Rafael M. Rosa
Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brazil. E-mail: Email: rafaelmassonrosa (at) usp (dot) br
https://doi.org/10.5281/zenodo.8226295
Zoo Tycoon 2 is a zoo-building simulation game developed by Blue Fang Games and originally released in 2004 as a sequel to the classic Zoo Tycoon game. In Zoo Tycoon 2 (henceforth referred to as ZT2), the player’s goal is to build a successful zoo, which involves not only creating an aesthetically pleasing design but also making sure it is able to fulfill the needs of both your guests and, most importantly, your animals. With animals as its central element, ZT2 offers the player dozens of different species, each with its own conservation status, enrichment needs and habitat preferences. The game puts great emphasis on environmental education, depicting the importance of zoos as tools in conservation and rewarding the player for fulfilling specific conservation goals, such as reintroducing individuals from endangered species to the wild. Additionally, the game even has its own in-game encyclopedia (dubbed Zoopedia) with additional information on each species’ taxonomy, ecology and conservation.
Over its development, ZT2 spawned four expansion packs adding new species and other content focused around a specific theme: Endangered Species, African Adventure, Marine Mania and Extinct Animals. In total, the game with all expansions has 130 animal species available to the player. It is also interesting to note that, since its release, ZT2 has nurtured a very prolific modding community that is still active over ten years since the game’s official development ceased. Over the years, several thousands of user-made mods were added to the game, ranging from simple additions to total remakes of the original game (ZT2 Download Library, 2021). As such, it is clear that ZT2 is still a much beloved game and also a powerful tool in representing biodiversity, as a fully modded game can have thousands[1] of real animal species from many different taxa.

This paper will focus on the game’s last official expansion pack, Extinct Animals. As hinted by its title, this expansion focuses on adding extinct species to the game. It contains a roster of 31 animals, ranging from long-lost creatures from millions of years ago to species that have only become extinct in the past decades. As a way to justify their inclusion in a zoo game, the expansion includes a minigame in which the player has to hunt for fossils from each species, which can then be assembled and used as a source of DNA for cloning them into your zoo. This is, of course, nonsense from a scientific point of view. DNA degrades quickly over time and the very few genetic samples we have from fossils are largely incomplete and only present in specimens from up to 1.5 million years ago (Willerslev et al., 2004; Kirkpatrick et al., 2016). Still, it is an interesting way to engage the player in paleontology and deserves praise for not outright copying the “fossil mosquitoes” idea from Jurassic Park.
As mentioned above, environmental education is one of the core aspects of the game. ZT2 attempts to present the animal species in a relatively accurate (albeit still very stylized) way. This is a no-brainer for extant species, as we all know how they live and what they look like. However, creating an accurate reconstruction is a challenge when it comes to extinct species. Even someone unfamiliar with paleontology will notice that every time you see a dinosaur depicted in a movie, videogame or toy, it will look different from other depictions of the same animal. This is not only due to artistic liberties taken by each media, but also because of the gaps in our knowledge of these animals. The fossil record, our main source of information on prehistoric organisms, only offers limited data and fossils are often fragmentary and incomplete. As such, the process of reconstructing prehistoric animals is always a work in progress, with new discoveries adding to our knowledge and prompting updated reconstructions. In this context, paleoart[3] plays an important role in translating paleontological knowledge to the greater public and inspiring people to learn more about these animals (Witton et al., 2014).
With this in mind, this paper reviews the paleontological reconstructions presented in Zoo Tycoon 2: Extinct Animals. Since the game contains too many species to discuss at once, this article focuses specifically on the dinosaurs (including birds), which are arguably the most popular group of extinct animals. An interesting detail about this is that the ZT2 not only depicts the animals’ appearances, but also aspects of their ecology. As such, alongside with their anatomy, it is possible to evaluate whether the ecological aspects presented in the game are accurate considering what we currently know about these animals.
A QUICK OVERVIEW OF DINOSAUR BIODIVERSITY
Dinosaurs are a very diverse and well-known group of animals. The first dinosaurs are thought to have evolved in the Late Triassic period, approximately 245 million years ago. They would then come to dominate most terrestrial ecosystems for the next two geological periods, the Jurassic and the Cretaceous, before facing a devastating mass extinction event at the end of the Cretaceous period. The Mesozoic era, comprising the Triassic, Jurassic and Cretaceous periods, has thus been dubbed “the age of reptiles” due to the predominance of dinosaurs and other large reptiles. Despite suffering many losses during the Cretaceous-Paleogene mass extinction event, the remaining dinosaurs (birds) quickly diversified and are still the most diverse group of terrestrial vertebrates in modern times (Benton, 2014; Sues, 2019).
There are approximately 1208 well-established genera of non-avian dinosaurs and several genera of extinct birds (Olshevsky, 2021). As the fossil record is largely incomplete, this number represents only a fraction of the total diversity of dinosaurs in the past. In addition, there are nearly 11,000 described species of birds alive today (Clements et al., 2019; Gill et al., 2021) and new dinosaur species (both extinct and living) are still discovered every year.
There are three major clades (or groups) of dinosaurs: Ornithischia, Sauropodomorpha and Theropoda. In recent years, there have been some controversies concerning the relationships between these clades (Baron et al., 2017; Langer et al., 2017). In spite of that, the classic and most widely adopted hypothesis proposes that theropods and sauropodomorphs are more closely related to each other than to ornithischians, with both Theropoda and Sauropodomorpha constituting a clade named Saurischia (Fig. 2).
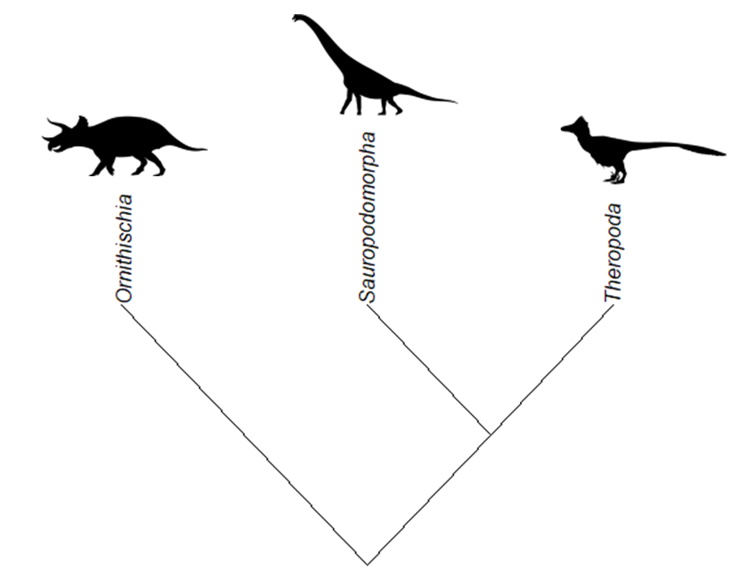
Understanding those relationships gives us some context as to where each species fits in the tree of life. It’s not this article’s intent to give a detailed account of dinosaur phylogeny and evolution, but this basic framework will be useful to discuss how this diversity is translated to ZT2.
DINOSAURS IN ZOO TYCOON 2: EXTINCT ANIMALS
Out of the three major clades of dinosaurs, only Ornithischia and Theropoda are represented in ZT2. This is unfortunate, since Sauropodomorpha is a very interesting lineage that includes some of the earliest dinosaurs and also some of the largest terrestrial animals of all time (Benton, 2014), so there are many iconic sauropodomorphs that could have been represented in the game. The ornithischians and theropods represented in ZT2 are discussed separately below.
Ornithischia
In total, five species of ornithischians are represented in ZT2: Ankylosaurus magniventris, Kentrosaurus aethiopicus, Stegosaurus stenops, Styracosaurus albertensis and Triceratops horridus. Out of these, A. magniventris, K. aethiopicus and Se. stenops belong to the clade Thyreophora, while Sy. albertensis and T. horridus belong to the clade Ceratopsia.
Ornithischians were one of the two major clades of herbivorous dinosaurs during the Mesozoic, the other being the sauropodomorphs. Among ornithischians, thyreophorans were distinctive for many of them possessed rows of plates, spikes or large scutes covering their backs, which led to them being often described as “armored” dinosaurs. Ceratopsians, in turn, are known for the large frills and horns that some of its later members bore on their heads (Benton, 2014; Sues, 2019). Those two clades include some of the most famous dinosaurs, but Ornithischia also includes other very iconic lineages, such as the pachycephalosaurs and the duck-billed dinosaurs, which sadly they are not represented in ZT2.

A common trend seen among ZT2’s ornithischians (and other animals as well) is that their proportions are a bit off. Their heads are usually too large, the legs are too stubby and the body is overall too short. This is likely due to the cartoon-like aesthetic adopted by the game and more of a case of stylization than of anatomical mistake. Ankylosaurus in particular seems to suffer from this (Fig. 3A), since the real animal was actually quite long and comparatively not very tall (Carpenter, 2004; Arbour & Mallon, 2017), while the ZT2 representation is much taller and not long enough. Moreover, the feet on all the ornithischians in ZT2 are too stocky and cylindrical, looking more like the feet of elephants than what would be expected of real ornithischians.
Odd proportions aside, there are relatively few anatomical inaccuracies in ZT2’s ornithischians. One of those is that the Kentrosaurus in ZT2 has its pair of lateral spikes placed on its hips (Fig. 3C). This was common in past reconstructions, but more modern reconstructions place these spikes on the animal’s shoulders, since close relatives of Kentrosaurus were known to have similar shoulder spikes (Galton & Upchurch, 2004). Another inaccurate detail is related to the fact that real-life Stegosaurus had its throat covered by an armor of ossicles (Fig. 4), which likely granted them protection from predators (Main et al., 2005). This is not represented in ZT2’s Stegosaurus (Fig. 3E), although it is a relatively minor detail that would likely be barely visible in the game.

It should be mentioned that the ornithischians in ZT2 are featherless and presumably covered in scales. Feather-like integument has been recovered in some early ornithischian fossils, including ceratopsians (Mayr et al., 2002; Zheng et al., 2009; Ksepka, 2020). As such, it is possible that such structures were present to some extent in other ornithischians. As a matter of fact, it has even been suggested that feathers or feather-like structures could be an ancestral feature of all dinosaurs and other related lineages (Yang et al.¸ 2019; Ksepka, 2020). However, current evidence suggests that the bodies of large ceratopsians and thyreophorans, such as the ones represented in ZT2, were covered mostly or entirely in scales (Ksepka, 2020). Thus, since we have no direct evidence of feathers in the species represented in ZT2, depicting them with a featherless skin is a valid choice.
The biome preferences of the ornithischians in ZT2 are a mixed bag. Both Ankylosaurus and Kentrosaurus are depicted as living in the wetlands biome. Data from the geological formations in which Ankylosaurus fossils were recovered indicate that it inhabited an environment similar to a subtropical or temperate forest (Johnson, 1997; Ősi et al., 2016). Kentrosaurus, in turn, is found in a formation that is associated with coastal lagoons, plains and vegetated inlands with a warm and dry climate (Aberhan et al., 2002). Thus, it is possible that wetlands were frequented by both Ankylosaurus and Kentrosaurus, but it seems that they are not a good representative of their primary habitats.
Stegosaurus and Triceratops, in turn, are depicted in ZT2 as having a preference for the boreal forest biome. This is likely a reference to the fact that gymnosperms, like conifers, were the most abundant plants during the Mesozoic, while today they are only dominant in a few ecosystems, such as boreal forests (Leslie et al., 2018; Condamine et al., 2020). However, the climate during the Jurassic and Cretaceous periods (when Stegosaurus and Triceratops lived, respectively) was far warmer than it is today, making cold environments like today’s boreal forests very rare or even absent (Nordt et al., 2003; Sellwood & Valdes, 2008; Huber et al., 2018). Stegosaurus fossils have been found in a formation associated with a semi-arid environment composed of forests and fern savannas (Chure et al., 2006). Triceratops fossils have been recovered from several distinct geological formations, most associated with a variety of subtropical environments (Estes & Berberian, 1970; Derstler, 1994). As such, the choice of attributing one of today’s coldest biomes to Stegosaurus and Triceratops is quite questionable.
Styracosaurus, on the other hand, is represented with a preference for tropical rainforests in ZT2. The formations in which Styracosaurus fossils have been found indicate a floodplain or swamp-like environment with a warm climate (Eberth, 2005). Tropical rainforest is overall not a bad choice to represent such environments, but perhaps wetlands would be a better choice. Nonetheless, it is important to note that biomes in the distant past were likely very different from any of today’s biomes, making it difficult to find accurate analogs for them.
Theropoda
In total, nine extinct species of theropods are represented in ZT2: Carnotaurus sastrei, Stokesosaurus clevelandi, Tyrannosaurus rex, Protarchaeopteryx robusta, Deinonychus antirrhopus, Utahraptor ostrommaysi, Velociraptor mongoliensis, Aepyornis maximus (the elephant bird) and Raphus cucullatus (the dodo). Out of these, C. sastrei belongs to the clade Abelisauridae; S. clevelandi and T. rex belong to the clade Tyrannosauroidea; P. robusta belongs to the clade Oviraptorosauria; D. antirrhopus, U. ostrommaysi and V. mongoliensis belong to the clade Dromaeosauridae; and A. maximus and R. cucullatus belong to the clade Aves.
Theropoda is the most diverse out of the three major clades of Dinosauria and includes most carnivorous dinosaurs, some of which were the largest land predators of all time, although many omnivorous and herbivorous theropods are also known (Benton, 2014; Sues, 2019). Only a few lineages of theropods are represented in ZT2. Carnotaurus is only distantly related to the other theropods mentioned here, being a representative of a strange lineage of theropods that were the main predators in the southern continents during the Cretaceous. The remaining theropods in ZT2 are part of a large clade known as Coelurosauria, which includes a wide variety of dinosaurs ranging from the T. rex to birds (Benton, 2014; Hendrickx, Hartman & Mateus, 2015; Sues, 2019). In fact, both A. maximus and R. cucullatus are coelurosaurs and the only extinct dinosaurs in ZT2 that did not live during the Mesozoic.

Overall, the theropods in ZT2 have similar problems to the ornithischians. Namely, their proportions are quite odd and cartoonish, with heads and feet being too large in comparison to the bodies. In large theropods, such as Carnotaurus and Tyrannosaurus, the back is often arched upwards (Fig. 5A, E), while it should be essentially straight and horizontal. Furthermore, the heads of both Carnotaurus and Deinonychus look a bit too blocky and only vaguely resemble their real shapes (Fig. 5A, I). Another mistake, shared by most non-avian theropods in ZT2, is that their hands are pronated, which means their palms are facing down. Generally speaking, theropods did not possess the ability to pronate their hands and instead had their palms facing each other constantly, with very limited movement (Carpenter, 2002).
When it comes to feathers, theropods in ZT2 are a mixed bag. Carnotaurus, Stokesosaurus, Tyrannosaurus and Deinonychus are depicted as featherless, while the remaining theropods are at least partially feathered. For Carnotaurus, this is actually fine, since its fossil contains extensive skin impressions that suggest it was covered by scales (Czerkas & Czerkas, 1997). In regards to the two tyrannosauroids, the situation is trickier. Feathers have been found on several early tyrannosauroids and close relatives, indicating that they were an ancestral trait of this lineage. However, skin impressions of Tyrannosaurus suggest that at least large portions of its body were featherless, which could be attributed to the evolution of gigantism in these animals (Bell et al., 2017). Although this matter is still debated, it seems that depicting Tyrannosaurus as featherless is fine. Stokesosaurus, on the other hand, is known only from a few bones (that is, no skin impressions are known). Considering it was a small-sized animal and closely related to other Jurassic tyrannosauroids, which are known to be feathered (Loewen et al., 2013), it’s quite likely that Stokesosaurus would also be covered in feathers.
As for the remaining theropods in ZT2, however, the presence of extensive feather coverings is almost certain. Dromaeosaurids, such as Velociraptor, are among the closest relatives of birds and some of the most unambiguously feathered dinosaurs we know of (Xu et al., 2003; Turner et al., 2007). While we still haven’t found direct evidence of feathers in Deinonychus and Utahraptor fossils, it is only safe to assume that they were as feathered as their close relatives. The featherless Deinonychus depicted in ZT2 (Fig. 5I) is likely inspired by popular depictions of dromaeosaurids, such as those of Jurassic Park, but this is particularly puzzling considering that Velociraptor itself is depicted as feathered. Utahraptor is also depicted as almost featherless (Fig. 5K), with only the male possessing a crest of feathers on its head, which is also not nearly enough to represent what the real animal would have looked like. In regards to Velociraptor, while it is certainly the most accurately feathered dromaeosaurid in the game (Fig. 5M), its arms are mostly featherless, which conflicts with the evidence of large feathers constituting bird-like wings on this animal (Turner et al., 2007).
Protarchaeopteryx is case of its own (Fig. 5G). In real life, it was a small and feathered oviraptorosaur known only from a single skeleton (Qiang et al., 1998). As an oviraptorosaur, it was more closely related to dinosaurs like Oviraptor and Incisivosaurus than to birds. In ZT2, however, Protarchaeopteryx is depicted in a much more bird-like fashion, looking essentially like a bird with long tail, hands and teeth. In fact, it seems that the ZT2 Protarchaeopteryx was heavily based on the game’s secretary bird, as it has a similar model and uses almost the same animations. While oviraptorosaurs certainly were bird-like, it seems like the game is trying to depict Protarchaeopteryx as an extinct bird like the much more famous and similar-named Archaeopteryx. This is further corroborated by the fact that the Protarchaeopteryx fossil used in the fossil-building minigame of ZT2 is clearly copied from the famous Berlin Archaeopteryx specimen (Fig. 6). Despite the similar names, Protarchaeopteryx and Archeopteryx are not closely related and not really similar at all. Perhaps the game developers intended to add Archaeopteryx to the game at first, but later decided to change it to a larger and more terrestrial animal to make its implementation easier, or maybe to draw attention to a more obscure and often overlooked species.

The two extinct birds depicted in ZT2 are overall more accurate than their non-avian relatives. Interestingly, the elephant bird depicted in ZT2 is said to be Aepyornis maximus, which was considered the largest species of elephant bird by the time of the game’s release. However, a recent study reclassified the largest A. maximus specimens in a new genus and species, Vorombe titan, which is now the largest species of elephant bird (Hansford & Turvey, 2018). It is uncertain whether the elephant bird in ZT2 should also be considered a representation of V. titan, since the game developers were likely trying to represent the largest elephant birds, or if it should remain A. maximus as was originally assigned. Regardless, its anatomy (Fig. 5O) is quite suitable and not so inaccurate for both Aepyornis and Vorombe, even though some details, such as the feather crest on its head, are merely speculative.
The dodo in ZT2 is also relatively accurate (Fig. 5P), but some things should be noted in regards to its color. Namely, it is depicted with a light blue and gray coloration. This is likely inspired by famous gray or blueish-gray depictions of the dodo, such as the Edwards’ Dodo painting by Dutch painter Roelant Savery (late 1620s; Fig. 7). However, the accuracy of such depictions has been questioned (Dissanayake, 2004). The most accurate depiction of a dodo is considered to be that of Mughal painter Ustad Mansur (c. 1625; Fig. 7), which shows the bird with a mostly brown plumage. The brown color of Mansur’s dodo is consistent with other contemporary descriptions of this bird, indicating that dodos likely had a brown hue (Hume, 2006). As such, the light blue hue depicted in ZT2 seems to be inaccurate. It should be noted that most times we have no evidence on the color of extinct species and this discussion is relevant to the dodo only because it became extinct in historical times and so, many contemporary paintings and descriptions are available.

As a side note, there is another bird species present in ZT2’s Extinct Animals expansion: the killer penguin. This is a fictional animal that can only be obtained by failing the cloning minigame. The killer penguin is depicted as a very large penguin with a toothed beak and eyes that glow. Overall, its design is similar to the rockhopper penguins of genus Eudyptes and the game even gives it the fictional scientific name Eudyptes omnicidus. It is depicted as very violent, being able and willing to kill literally every other animal in the game. The killer penguin in ZT2 is an easter egg and likely a reference to a bug in the original Zoo Tycoon game that made the emperor penguin able to kill most other animals. It is also described as being native to Madagascar, which is probably a reference to the penguins in the Madagascar movie franchise. Nonetheless, despite the killer penguin being a fictional species, it is interesting to note that giant penguin species are known to have lived in the early Cenozoic era (Fig. 8) and some, such as Anthropornis nordenskjoldi, even reached sizes comparable to adult humans (Jadwiszczak, 2001).

Just like with the ornithischians, the biome preferences of the theropods in ZT2 contain a few odd choices. Carnotaurus, Stokesosaurus and Utahraptor are represented as boreal forest animals in-game, which is very unlikely for the reasons discussed earlier. The geological formation in which Carnotaurus was found is associated with an environment composed of estuaries and coastal plains (Pascual et al., 2000), although it is possible that it lived in other environments as well. Stokesosaurus was found in the same formation as Stegosaurus and thus probably shared the same semi-arid environment (Chure et al., 2006). Utahraptor was also found in a formation associated with a semi-arid environment composed of prairies and open woodlands (Joeckel et al., 2019).
Tyrannosaurus, Deinonychus, the elephant bird and the dodo are represented as tropical rainforest animals in ZT2. Tyrannosaurus fossils have been found in many different geological formations with paleoenvironments including subtropical plains and forests, both humid and semi-arid (Estes & Berberian, 1970; Derstler, 1994; Jasinski et al., 2011), indicating it could tolerate a wide range of habitats. Deinonychus has been found in formations indicating an environment composed of tropical or subtropical forests and swamps (Forster, 1984; Norell & Makovicky, 2004; Wedel & Cifelli, 2005), thus tropical rainforest is a suitable choice, although the wetlands biome would arguably be more accurate. The elephant bird, more specifically Aepyornis maximus, appears to have lived in forested environments in Madagascar (Torres & Clarke, 2018), making a tropical rainforest a reasonable choice as well. It is thought that the dodo lived in coastal woodlands in Mauritius (Fuller, 2002), which also makes tropical rainforest a relatively accurate choice.
Protarchaeopteryx is associated with the wetlands biome in-game, but the formation in which its fossils were found indicates an environment more akin to a temperate forest (Zhou, 2006). Velociraptor is associated with the desert biome, which is accurate, since the geological formation in which its fossils were found indicates a hot arid environment covered by sand dunes (Dashzeveg et al., 2005).
NOT A DINOSAUR, BUT A CLOSE RELATIVE
Also included in ZT2, Deinosuchus hatcheri (Fig. 9) is an extinct species of crocodilian that lived in North America during the Late Cretaceous period (Cossete & Brochu, 2020). Despite being often compared to them, crocodilians are not dinosaurs. However, both crocodilians and dinosaurs are grouped in a large clade named Archosauria, which also includes pterosaurs and other lesser-known extinct reptiles.
In modern times, crocodilians are the closest living relatives to birds, which also makes them the closest living relatives to dinosaurs as a whole (Benton, 2014; Grigg, 2015; Sues, 2019). As such, while Deinosuchus is not a dinosaur, it is part of a closely related lineage that shared its environment with many dinosaur species, making it appropriate to have a few comments about it here.

In regards to its anatomy, the ZT2 Deinosuchus is a bit strange (Fig. 9). Deinosuchus is a close relative of species in the genus Alligator and so, it probably looked somewhat similar to today’s alligators. The overall shape of ZT2’s Deinosuchus is certainly alligator-like, although its proportions are also exaggerated and it could be argued that its snout is too short and narrow. The skull of Deinosuchus was proportionally larger, broader and longer than that of Alligator, with its dorsal profile being almost rectangular (Fig. 10). A strange detail about the Deinosuchus in ZT2 is that its skin is covered in what look like spike-like osteoderms[4] (Fig. 9). The osteoderms on the real Deinosuchus were of similar shape to those of modern crocodilians and certainly not spiky like the ones in ZT2 (Cossete & Brochu, 2020).

Deinosuchus is represented with a preference for the wetlands biome in ZT2. This is relatively accurate, as the real Deinosuchus likely spent most of its time inside or nearby water and wetlands is the only primarily freshwater biome in ZT2. However, fossils of Deinosuchus are often found in formations associated with estuaries and brackish water bays, indicating that these animals might have inhabited coastal environments as well (Anglen & Lehman, 2000; Schwimmer, 2002).
CONCLUSION
Out of fourteen dinosaur species presented in Zoo Tycoon 2: Extinct Animals, five are ornithischians and nine are theropods. Since theropods are the most diverse lineage of dinosaurs, it is understandable that they have the largest number of species in-game as well. The species roster in ZT2 is interesting, as it includes not only famous dinosaurs, such as Tyrannosaurus and Triceratops, but also some rather obscure and lesser-known animals, like Stokesosaurus and Protarchaeopteryx. As such, the game serves its purpose as an educational tool by representing species that probably would not have reached the public’s attention otherwise. However, the absence of sauropodomorphs in ZT2 is a wasted potential, as it leaves out one of the major clades of dinosaurs; its inclusion could have been an opportunity to represent some of the largest animals that ever walked the Earth.
Overall, it seems that the ornithischians are relatively more accurate than the theropods in ZT2. However, although there are many inaccuracies in the game’s portrayals of the dinosaurs, it can be said that most of the time it does a good job at representing the most distinctive features of each species. The representation of ecological aspects for each species, such as their biome preferences, offers an interesting opportunity to discuss paleoecology as well, while most discussions regarding the accuracy of dinosaur reconstructions are purely restricted to anatomy.
REFERENCES
Aberhan, M.; Bussert, R.; Heinrich, W.; Schrank, E.; Schultka, S.; Sames, B.; et al. (2002) Palaeoecology and depositional environments of the Tendaguru Beds (Late Jurassic to Early Cretaceous, Tanzania). Fossil Record 5: 19–44.
Anglen, J.J. & Lehman, T.M. (2000) Habitat of the giant crocodilian Deinosuchus, Aguja Formation (Upper Cretaceous), Big Bend National Park, Texas. Journal of Vertebrate Paleontology 20: 26A.
Arbour, V.M. & Mallon, J.C. (2017) Unusual cranial and postcranial anatomy in the archetypal ankylosaur Ankylosaurus magniventris. FACETS 2: 764–794.
Baron, M.; Norman, D.; Barrett, P. (2017) A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 543: 501–506.
Bell, P.R.; Campione, N.E.; Persons, W.S.; Currie, P.J.; Larson, P.L.; Tanke, D.H.; Bakker, R.T. (2017) Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution. Biology Letters 13: 20170092.
Benton, M.J. (2014) Vertebrate Palaeontology. 4th Ed. John Wiley & Sons, Chichester.
Carpenter, K. (2002) Forelimb biomechanics of non-avian theropod dinosaurs in predation. Senckenbergiana Lethaea 82: 59–76.
Carpenter, K. (2004) Redescription of Ankylosaurus magniventris Brown 1908 (Ankylosauridae) from the Upper Cretaceous of the Western Interior of North America. Canadian Journal of Earth Sciences 41: 961–986.
Chure, D.J.; Litwin, R.; Hasiotis, S.T.; Evanoff, E.; Carpenter, K. (2006) The fauna and flora of the Morrison Formation: 2006. New Mexico Museum of Natural History and Science Bulletin, Albuquerque 36: 233–249.
Clements, J.F.; Schulenberg, T.S.; Iliff, M.J.; Billerman, S.M.; Fredericks, T.A.; Sullivan, B.L.; Wood, C.L. (2019) The eBird/Clements Checklist of Birds of the World: v2019. Cornell University. Available from: https://www.birds.cornell.edu/clementschecklist/download/ (Date of access: 29/Dec/2021).
Condamine, F.L.; Silvestro, D.; Koppelhus, E.B.; Antonelli, A. (2020) The rise of angiosperms pushed conifers to decline during global cooling. Proceedings of the National Academy of Sciences 117: 28867–28875.
Cossette, A.P. & Brochu, C.A. (2020) A systematic review of the giant alligatoroid Deinosuchus from the Campanian of North America and its implications for the relationships at the root of Crocodylia. Journal of Vertebrate Paleontology 40: e1767638.
Czerkas, S.A. & Czerkas, S.J. (1997) The Integument and Life Restoration of Carnotaurus. In: Wolberg, D.I.; Stump, E.; Rosenberg, G.D. (Eds.) Dinofest International. Academy of Natural Sciences, Philadelphia. Pp. 155–158.
Dashzeveg, D.; Dingus, L.; Loope, D.B.; Swisher III, C.C.; Dulam, T.; Sweeney, M.R. (2005) New stratigraphic subdivision, depositional environment, and age estimate for the Upper Cretaceous Djadokhta Formation, southern Ulan Nur Basin, Mongolia. American Museum Novitates 3498: 1−31.
Derstler, K. (1994) Dinosaurs of the Lance Formation in Eastern Wyoming. In: Nelson, G.E. (Ed.). The Dinosaurs of Wyoming. 44th Annual Field Conference Guidebook. Wyoming Geological Association, Casper. Pp. 127–146.
Dissanayake, R. (2004) What did the dodo look like? The Biologist 51: 165–168.
Eberth, D.A. (2005) The Geology. In: Currie, P.J. & Koppelhus, E.B. (Eds.) Dinosaur Provincial Park: a spectacular ancient ecosystem revealed. Indiana University Press, Bloomington. Pp. 54–82.
Estes, R. & Berberian, P. (1970) Paleoecology of a late Cretaceous vertebrate community from Montana. Breviora 343: 1–35.
Fordyce, R.E. & Ksepka, D.T. (2012) The strangest bird. Scientific American 307: 56–61.
Forster, C.A. (1984) The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana. The Mosasaur 2: 151–163.
Fuller, E. (2002) Dodo – From Extinction to Icon. HarperCollins, London.
Galton, P.M. & Upchurch, P. (2004) Stegosauria. In: Weishampel, D.B.; Dodson, P.; Osmólska, H. (Eds.) The Dinosauria. 2nd Ed. University of California Press, Berkeley. Pp. 343–362.
Gill, F.; Donsker, D.; Rasmussen, P. (2021) IOC World Bird List (v11.1). Available from: https://www.worldbirdnames.org/new/ioc-lists/crossref/ (Date of access: 29/Dec/2021).
Grigg, G. (2015) Biology and Evolution of Crocodylians. Cornell University Press, Ithaca.
Hansford, J.P. & Turvey, S.T. (2018) Unexpected diversity within the extinct elephant birds (Aves: Aepyornithidae) and a new identity for the world’s largest bird. Royal Society Open Science 5: 181295.
Hendrickx, C.; Hartman, S.A.; Mateus, O. (2015) An overview of non-avian theropod discoveries and classification. PalArch’s Journal of Vertebrate Palaeontology 12: 1–73.
Huber, B.T.; MacLeod, K.G.; Watkins, D.K.; Coffin, M.F. (2018) The rise and fall of the Cretaceous Hot Greenhouse climate. Global and Planetary Change 167: 1–23.
Hume, J.P. (2006) The history of the dodo Raphus cucullatus and the penguin of Mauritius. Historical Biology 18: 69–93.
Jadwiszczak, P. (2001) Body size of Eocene Antarctic penguins. Polish Polar Research: 147–158.
Jasinski, S.E.; Sullivan, R.M.; Lucas, S.G. (2011) Taxonomic composition of the Alamo Wash local fauna from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member) San Juan Basin, New Mexico. New Mexico Museum of Natural History and Science, Bulletin 53: 216–271.
Joeckel, R.M.; Ludvigson, G.; Moeller, A.; Hotton, C.L.; Suarez, M.B.; Suarez, C.A.; et al. (2019) Chronostratigraphy and terrestrial palaeoclimatology of Berriasian–Hauterivian strata of the Cedar Mountain Formation, Utah, USA. Geological Society London, Special Publications 498: 75–100.
Johnson, K.R. (1997) Hell Creek Flora. In: Currie, P.J. & Padian, K. (Eds.) The Encyclopedia of Dinosaurs. Academic Press, London. Pp. 300–02.
Kirkpatrick, J.B.; Walsh, E.A.; D’Hondt, S. (2016) Fossil DNA persistence and decay in marine sediment over hundred-thousand year to million-year time scales. Geology 44: 615–618.
Ksepka, D.T. (2020) Feathered dinosaurs. Current Biology 30: R1347–R1353.
Ksepka, D.T.; Betelli, S.; Giannini, N.P. (2006) The phylogeny of the living and fossil Sphenisciformes (penguins). Cladistics 22: 412–441.
Langer, M.C.; Ezcurra, M.D.; Rauhut, O.W.M.; Benton, M.J.; Knoll, F.; McPhee, B.W.; et al. (2017) Untangling the dinosaur family tree. Nature 551: E1–E3.
Leslie, A.B.; Beaulieu, J.; Holman, G.; Campbell, C.S.; Mei, W.; Raubeson, L.R.; Mathews, S. (2018) An overview of extant conifer evolution from the perspective of the fossil record. American Journal of Botany 105: 1531–1544.
Loewen, M.A.; Irmis, R.B.; Sertich, J.J.W.; Currie, P.J.; Sampson, S.D.; Evans, D.C. (2013) Tyrant dinosaur evolution tracks the rise and fall of Late Cretaceous oceans. PLOS ONE 8: e79420.
Main, R.P.; de Ricqlès, A.; Horner, J.R.; Padian, K. (2005) The evolution and function of thyreophoran dinosaur scutes: implications for plate function in stegosaurs. Paleobiology 31: 291–314.
Nordt, L.; Atchley, S.; Dworkin, S. (2003) Terrestrial evidence for two greenhouse events in the Latest Cretaceous. GSA Today 13: 4–9.
Norell, M.A. & Makovicky, P.J. (2004) Dromaeosauridae. In: Weishampel, D.B.; Dodson, P.; Osmólska, H. (Eds.) The Dinosauria. 2nd Ed. University of California Press, Berkeley. Pp. 196–210.
Olshevsky, G. (2021) Dinosaur Genera List. Available from: https://www.polychora.com/dinolist.html (Date of access: 29/Dec/2021).
Ősi, A.; Prondvai, E.; Mallon, J.; Bodor, E.R. (2016) Diversity and convergences in the evolution of feeding adaptations in ankylosaurs (Dinosauria: Ornithischia). Historical Biology 29: 539–570.
Pascual, R.; Goin, F.J.; González, P.; Ardolino, A.; Puerta, P.F. (2000) A highly derived docodont from the Patagonian Late Cretaceous: evolutionary implications for Gondwanan mammals. Geodiversitas 22: 395–414.
Qiang, J.; Currie, P.J.; Norell, M.A.; Shu-An, J. (1998) Two feathered dinosaurs from northeastern China. Nature 393: 753–761.
Schwimmer, D.R. (2002) Deinosuchus Localities and Their Ancient Environments. In: Schwimmer, D.R. (Ed.) King of the Crocodylians: The Paleobiology of Deinosuchus. Indiana University Press, Bloomington. Pp. 81–106.
Sellwood, B.W. & Valdes, P.J. (2008) Jurassic climates. Proceedings of the Geologists’ Association 119: 5–17.
Sues, H.D. (2019) The Rise of Reptiles: 320 million years of evolution. Johns Hopkins University Press, Baltimore.
Torres, C.R. & Clarke, J.A. (2018) Nocturnal giants: evolution of the sensory ecology in elephant birds and other palaeognaths inferred from digital brain reconstructions. Proceedings of the Royal Society B 285: 20181540.
Turner, A.H.; Makovicky, P.J.; Norell, M.A. (2007) Feather quill knobs in the dinosaur Velociraptor. Science 317: 1721.
Wedel, M.J. & Cifelli, R.L. (2005) Sauroposeidon: Oklahoma’s native giant. Oklahoma Geology Notes 65: 40–57.
Willerslev, E.; Hansen, A.J.; Rønn, R.; Brand, T.B.; Barnes, I.; Wiuf, C.; et al. (2004) Long-term persistence of bacterial DNA. Current Biology 14: R9–R10.
Witton, M., Naish, D.; Conway, J. (2014) State of the Palaeoart. Palaeontologia Electronica 17: 5E.
Xu, X.; Zhou, Z.; Wang, X.; Kuang, X.; Zhang, F.; Du, X. (2003) Four-winged dinosaurs from China. Nature 421: 335–340.
Yang, Z.; Jiang, B.; McNamara, M.E.; Kearns, S.L.; Pittman, M.; Kaye, T.G.; et al. (2019) Pterosaur integumentary structures with complex feather-like branching. Nature Ecology & Evolution 3: 24–30.
Zhou, Z. (2006) Evolutionary radiation of the Jehol biota: chronological and ecological perspectives. Geological Journal 41: 377–393.
ZT2 Download Library. (2021) ZT2 Download Library, an extensive library of user-made content and downloads for the videogame Zoo Tycoon 2. Available from: https://zt2downloadlibrary.fandom.com/wiki/ZT2_Download_Library_Wiki (Date of access: 29/Dec/2021).
About the author
Rafael Rosa is a biology undergraduate student at the University of São Paulo, Ribeirão Preto, who is very interested in zoology and paleontology. He is very fond of the Zoo Tycoon series and has played it since his childhood, which might have been one of the influences for getting him into biology in the first place.
[1] The game engine actually has a limit in how many assets can be added to the game before it crashes, so technically it can only hold a few hundred mods at the same time. Still, the player can pick and mix the mods they wish to have in a specific game session from the large pool of user-made content.
[2] As all other ZT2 images used herein. Archived at: https://web.archive.org/web/20121012035800/http://zootycoon.com/About/about_zoo_tycoon_2.htm
[3] The general term for artistic depictions of extinct animals.
[4] Osteoderms are bony deposits that form scales, plates or other structures on the skin of non-mammalian land vertebrates.