Skip to content
RSS Feed
Bluesky
Type your email…
Subscribe
Search
Journal of Geek Studies
About
Guidelines for authors
Current Issue
Archives
Vol. 13(1): Jun/2026
Vol. 12(2): Dec/2025
Vol. 12(1): Jun/2025
Vol. 11(2): Dec/2024
Vol. 11(1): Jun/2024
Older issues
Editorial Team
Partners
Contact
Behaviour
April 16, 2026
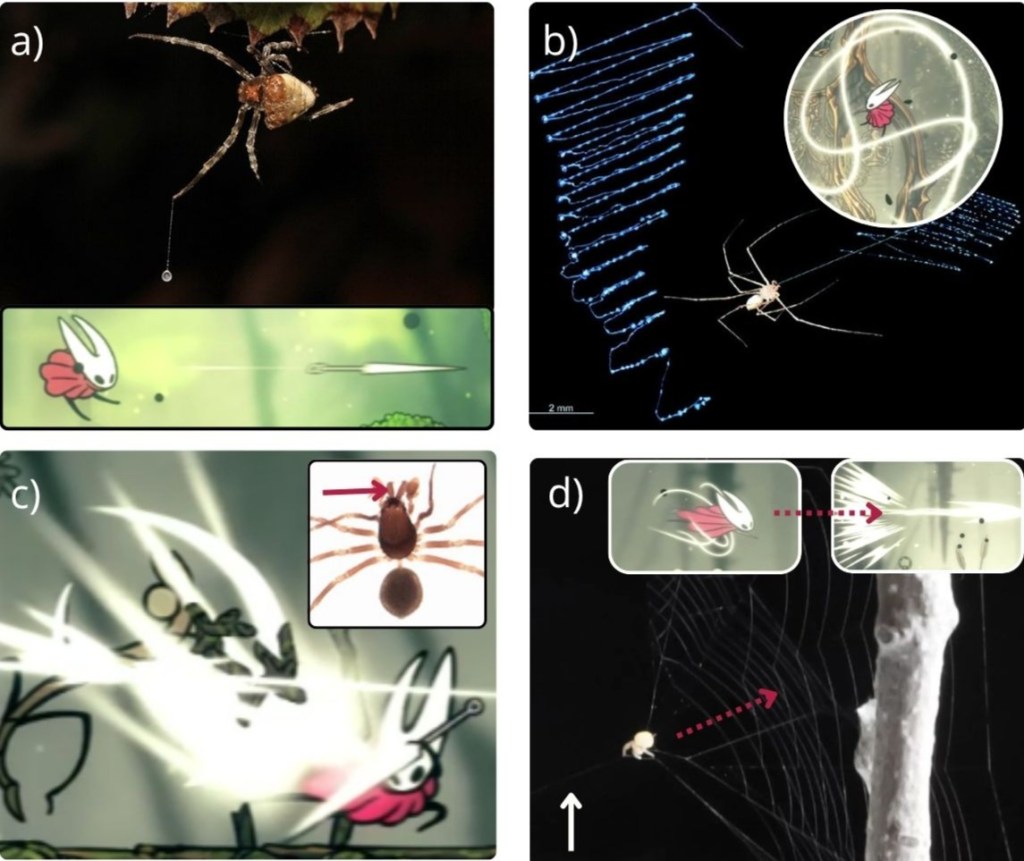
Hornet’s web: weaving the biology of silk into Hollow Knight: Silksong
Biology
,
Video Games
,
Zoology
Arachnology
,
Behaviour
,
Ecology
,
Hollow Knight
,
Metroidvania
February 15, 2022
The sea slugs of Shiroi Suna no Aquatope
Anime & Manga
,
Biology
Behaviour
,
Ecology
,
Malacology
,
Marine Biology
,
Taxonomy
,
Zoology
October 17, 2019
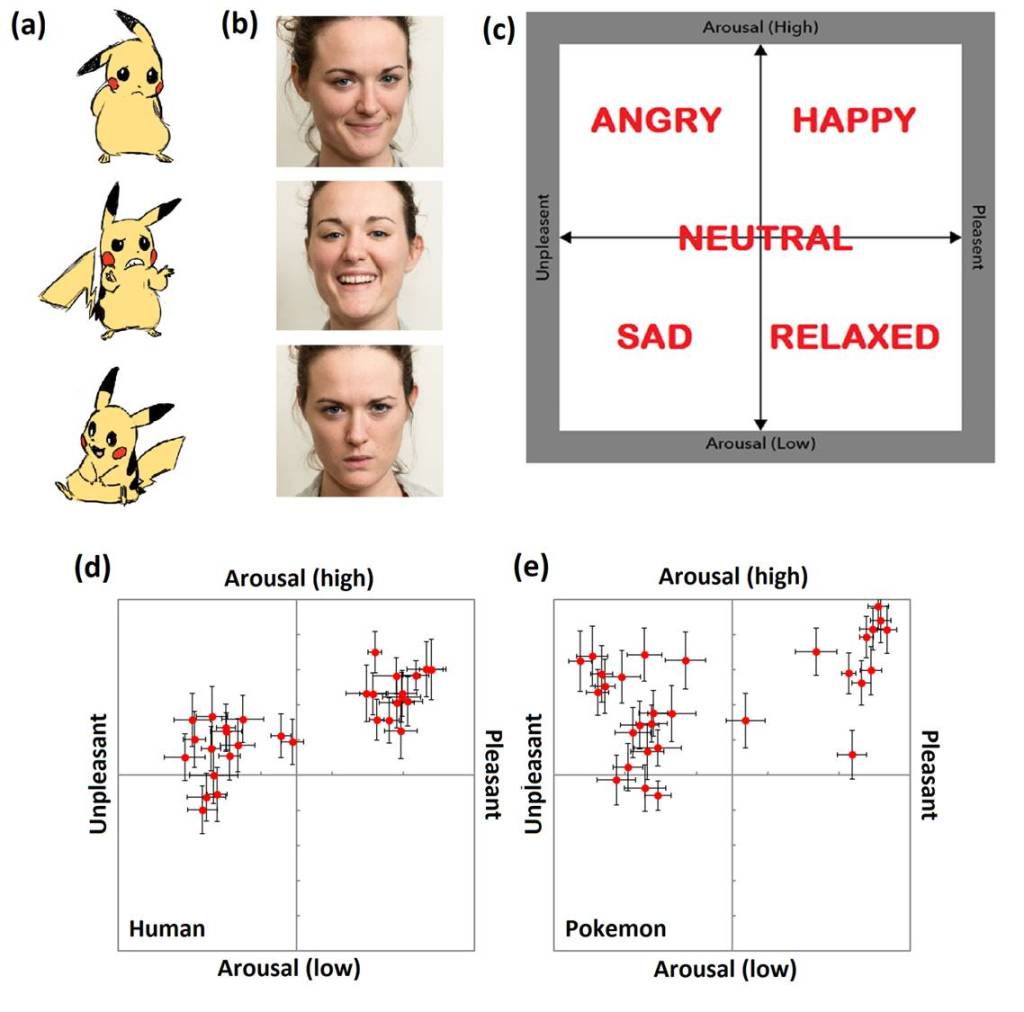
Perceiving the emotions of Pokémon
Anime & Manga
,
Psychology
,
Video Games
Behaviour
,
Pokémon
,
Psychophysics
October 17, 2018
Douglas Adams and the world’s largest, fattest and least-able-to-fly parrot
Biology
,
Literature
,
Zoology
Behaviour
,
Douglas Adams
,
Ecology
,
Ornithology
,
Zoology
September 18, 2016
Why (and how) Superman hides behind glasses: the difficulties of face matching
Comics
,
Psychology
,
Super Heroes
,
Television Series
Behaviour
,
DC Comics
,
Superman
Subscribe
Subscribed
Journal of Geek Studies
Join 305 other subscribers
Sign me up
Already have a WordPress.com account?
Log in now.
Journal of Geek Studies
Subscribe
Subscribed
Sign up
Log in
Report this content
View site in Reader
Manage subscriptions
Collapse this bar